Глава 1: Вычисление и мозг
Происхождение
Абиогенез
Спонтанное возникновение птиц и рыб из дерева, в зависимости от того, куда упадут его семена; Клод Дюре, Histoire admirable des plantes et herbes esmerueillables et miraculeuses en nature , 1605 Как началась жизнь на Земле? В девятнадцатом веке этот вопрос казался неразрешимым. Исследователи, пытавшиеся поймать жизнь в момент «спонтанного возникновения», остались ни с чем, как только научились дезинфицировать свои образцы. Как сказал Уильям Томсон (1824–1907), будущий лорд Кельвин, в своем выступлении перед Британской ассоциацией содействия науке в 1871 году: «Мёртвая материя не может стать живой, не подвергнувшись влиянию ранее живой материи. Это кажется мне таким же неоспоримым утверждением науки, как закон гравитации.» Существовала ли жизнь каким-то образом всегда? Пришла ли она на Землю из космоса, перенесённая астероидом? Томсон так думал, и некоторые до сих пор придерживаются этого мнения. Однако гипотеза «панспермии» не даёт нам ясного ответа. Откуда пришёл астероид и как жизнь возникла там ? Несмотря на чёткое изложение принципа эволюции, Чарльз Дарвин (1809–1882) тоже не имел представления. Поэтому в 1863 году он написал своему близкому другу Джозефу Долтону Хукеру, что «думать о происхождении жизни сейчас — это просто чепуха; можно так же хорошо думать о происхождении материи. » Сегодня у нас есть больше подсказок, хотя детали могут навсегда остаться потерянными в глубинах времени. Биологи и химики, работающие в области абиогенеза— изучения момента, когда, миллиарды лет назад, химия стала жизнью— разработали несколько правдоподобных историй о происхождении. В одной из них протобиологические организмы в древнем «мирe РНК» были организованы вокруг молекул РНК, которые могли как реплицироваться, так и сворачиваться в трехмерные структуры, способные действовать как примитивные ферменты.
В альтернативном подходе «метаболизм прежде всего» жизнь началась без генов, вероятно, в каменных трубках «черных курильщиков» на дне океана; РНК и ДНК появились позже. Возможно, в конечном итоге удастся опровергнуть одну из этих теорий... а может, и нет.
Мы вернемся к РНК и репликации, но давайте начнем с подробного рассмотрения версии, основанной на метаболизме, так как она проливает свет на проблему, которая ставила в тупик Дарвина и его последователей: как эволюция может начаться без генетической наследственности. Как мы увидим, абиогенез становится менее загадочным, когда мы развиваем более общее понимание эволюции — такого, которое может охватывать время до появления жизни.
▶ Лавовое озеро, Килауэа, Гавайи
Давайте вспомним о происхождении нашей планеты. Хадейская эра началась 4,6 миллиарда лет назад, когда Земля впервые конденсировалась из аккреционного диска каменных материалов, вращающегося вокруг нашей новорожденной звезды, Солнца. Мантия нашей планеты была более текучей, чем сегодня, и насыщенной горячими, недолговечными радиоактивными элементами, бурлила, выделяя углекислый газ и водяной пар. Поверхность представляла собой вулканический ад, светившийся озерами раскаленной лавы и изрытый серными вентиляционными отверстиями, извергающими дым.
Потребовались сотни миллионов лет, чтобы условия на поверхности стабилизировались. Согласно Международному астрономическому союзу, объект, вращающийся вокруг звезды, может считаться планетой только в том случае, если он имеет достаточную массу, чтобы либо поглотить, либо, проходя мимо, выбросить других обитателей из своей орбиты. Но в хаосе ранней Солнечной системы это заняло много времени. Пока Земля очищала свою орбиту от обломков, она постоянно подвергалась бомбардировке кометами и астероидами, включая гигантские ударные тела диаметром более шестидесяти миль. Один такой удар мог бы разогреть и без того удушающую атмосферу до тысячи градусов по Фаренгейту.
4-миллиардолетние детритовые цирконы из гадайских метаседиментов холмов Джек, террейн Нэрриер Гнейс, Западная Австралия
Одна из этих коллизий, похоже, произошла с другой вновь образовавшейся планетой, что едва не привело к полному уничтожению Земли. Согласно этой теории, достаточно разрушенной, расплавленной и испарившейся породы было выброшено на орбиту, чтобы сформировать Луну.
Неудивительно, что очень мало геологического материала сохранилось с гадайского периода — в основном это кристаллы циркона, заключенные в метаморфическом песчанике в холмах Джек, Западная Австралия. Трудно представить, что при таких суровых условиях могла возникнуть или выжить какая-либо жизнь. Возможно, каким-то образом, это все же произошло. Мы точно знаем, что к более позднему гадайскому или раннему архейскому периодам — не позже 3,6 миллиарда лет назад — поверхность остыла достаточно, чтобы конденсировался жидкий океан и началась химия жизни.
Сегодня эти ранние условия наиболее близко воспроизводятся черными курильщиками. Они образуются вокруг гидротермальных источников на срединноокеанских хребтах, где тектонические плиты расходятся, и формируется новая кора. В таких местах морская вода, просачиваясь в породу, контактирует с горячим магмой. Перегретая вода затем поднимается обратно, унося водород, углекислый газ и соединения серы, которые осаждаются, образуя курильщики под водой. Зонд, отправленный на тысячи футов вниз для исследования этих неземных сред, находит их полными странных форм жизни всех видов, привлеченных теплом, питательными веществами и друг другом. Некоторые из обитателей могут иметь очень глубокие корни в прошлом.
▶ «Канделябр» — черный курильщик в гидротермальном поле Логачева на Среднеатлантическом хребте, на глубине 10,800 футов
Как и лавовая порода, курильщики пористые, а поры представляют собой идеальные маленькие камеры для протекания химических реакций — потенциально питаемых удобным источником энергии, поскольку водородный газ создает протонные градиенты через тонкие стенки между порами, заставляя их работать как батареи.
Учитывая энергию, минералы железистого сульфида в горных породах, действующие как катализаторы, и углекислый газ, пузырящийся в растворе, самоподдерживающиеся цепочки химических реакций могут вспыхнуть, как мотор, который запускается. Возможно, именно это и было изначальным мотором жизни: примитивный, но почти стабильный метаболизм, еще без генов, ферментов или четко определенной границы между внутренним и внешним. Однако такие усовершенствования могли последовать довольно быстро, ведь этот химический синтетический мотор напоминает «обратный цикл Кребса», который сейчас широко считается источником энергии для самых ранних клеток.
Обычный или «прямой» цикл Кребса был открыт в 1937 году выдающимся биохимиком Гансом Кребсом (1900–1981). Скорее похожий на генератор, работающий на газе, чем на мотор, цикл Кребса лежит в основе того, как все аэробные организмы на Земле «сжигают» органическое топливо для высвобождения энергии, процесс, известный как «дыхание».
Входными данными для этого цикла химических реакций являются сложные органические молекулы (которые мы потребляем) и кислород (который мы вдыхаем); «выброс» включает углекислый газ и воду (которые мы выдыхаем). Вырабатываемая энергия поддерживает протонные градиенты через сложенные мембраны внутри наших митохондрий, а поток этих протонов затем питает все остальные клеточные функции.
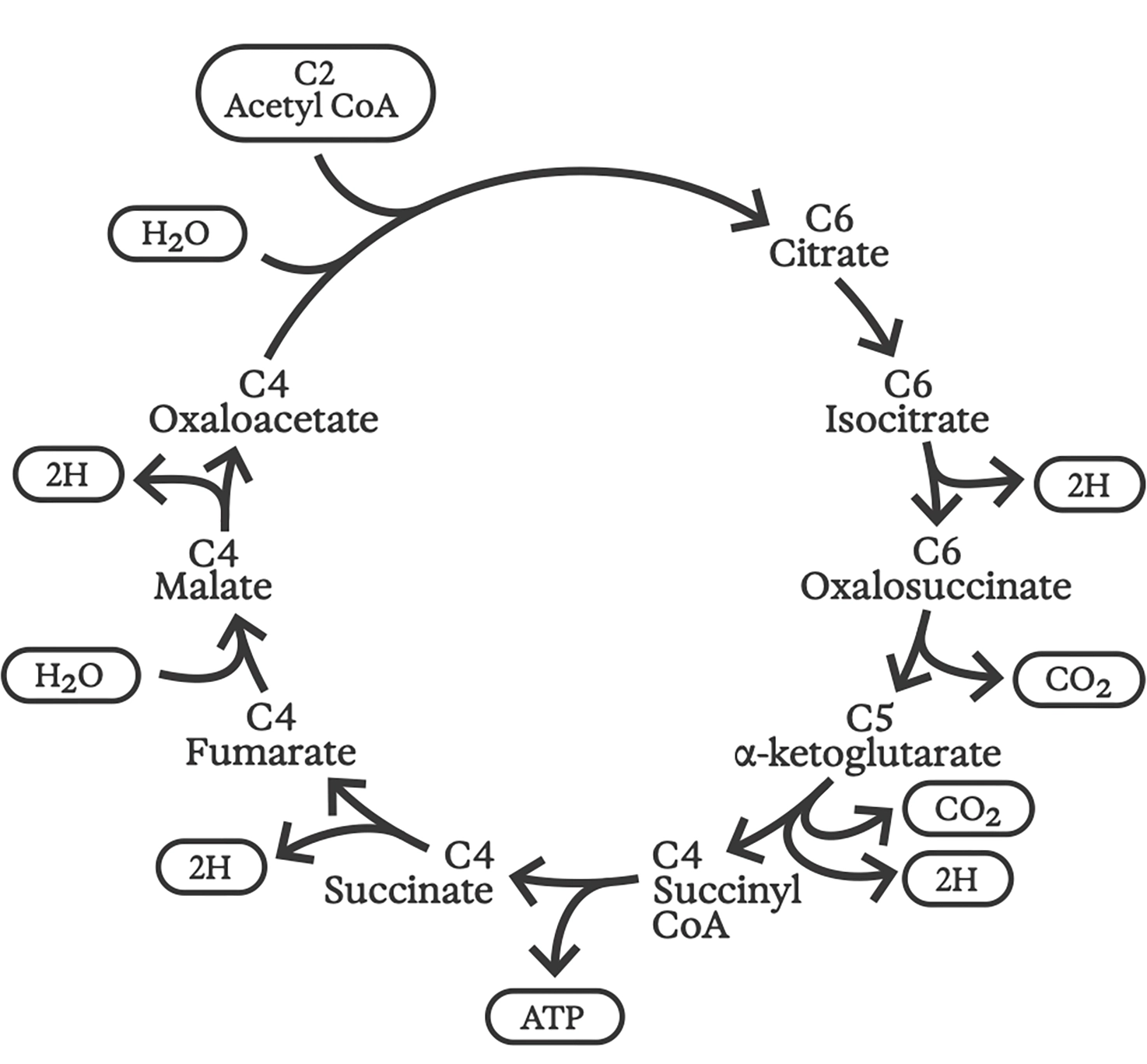
Цикл Кребса
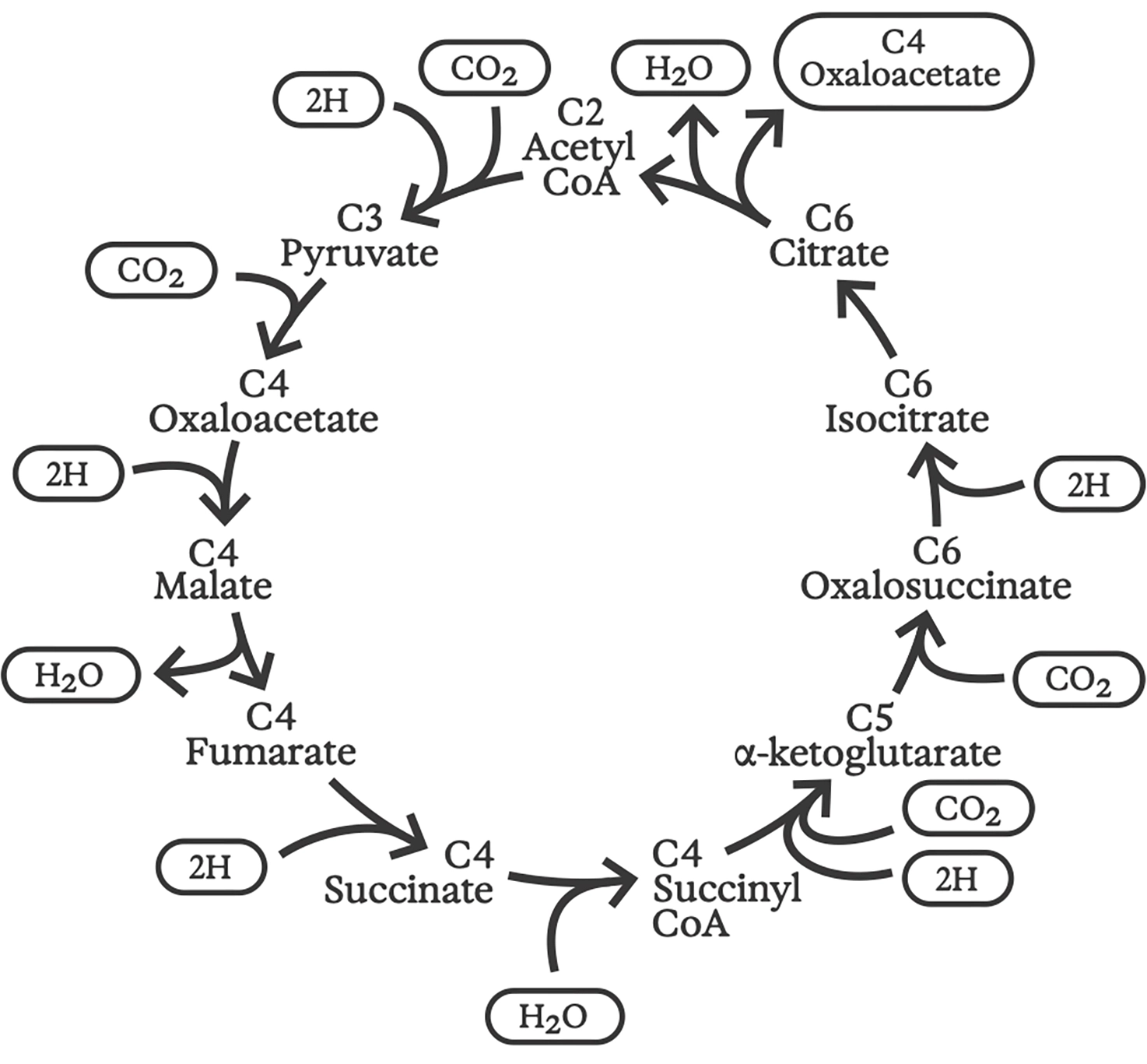
Обратный цикл Кребса
Идея «обратного цикла Кребса» была впервые предложена тройкой исследователей из Университета Калифорнии в Беркли в 1966 году. Она оставалась спорной на протяжении десятилетий, но теперь известно, что этот цикл обеспечивает фиксацию углерода в древних анаэробных серобактериях — некоторые из которых до сих пор обитают в глубоководных гидротермальных источниках. Как следует из названия, обратный цикл Кребса состоит примерно из тех же химических реакций, что и прямой цикл, но протекает в противоположном направлении.
Начав с воды и углекислого газа, протонные градиенты приводят к синтезу всех основных строительных блоков, необходимых для клеточной структуры и функции, включая сахара, аминокислоты для белков, жирные кислоты и изопрены для клеточных мембран, а также нуклеотиды для построения РНК и ДНК.

Рентгеновский снимок русских матрешек
Вся жизнь произошла из моря, и внутреннее пространство каждой клетки каждого живого организма воспроизводит ту соленую, водянистую среду — крошечный «океан внутри». Это общеизвестный факт. Но наши митохондрии, «электростанции» внутри клеток, где происходит дыхание, возможно, на самом деле воспроизводят гораздо более специфическую глубоководную химию черного курильщика. Почти в буквальном смысле наши тела похожи на русские матрешки — мембраны внутри мембран, и каждая вложенная среда воссоздает более раннюю стадию эволюции жизни на Земле. Чем глубже мы заглядываем в себя, тем дальше можем увидеть в прошлое. Прекрасная, волнующая мысль.
Симбиогенез
Неважно, что появилось первым — РНК или метаболизм, даже самые простые бактерии, выживающие сегодня, являются результатом множества последующих эволюционных шагов. Однако, в отличие от повседневной, постепенной мутации и отбора, которые представлял себе Дарвин, самые важные из этих шагов могли быть крупными и резкими. Эти «крупные эволюционные переходы» связаны с тем, что более простые, менее сложные реплицирующие сущности становятся взаимозависимыми, образуя более крупный, более сложный и более способный репликатор.
▶
Трансмиссионная электронная микроскопия, показывающая митохондрии в клетках
Как выяснила независимый биолог Линн Маргулис (1938–2011) в 1960-х годах, эукариотические клетки, такие как те, что составляют наши тела, являются результатом такого события.
Приблизительно два миллиарда лет назад бактерии, ставшие нашими митохондриями, были поглощены другой одноклеточной формой жизни, похожей на современные археи — крошечные, столь же древние микроорганизмы, которые продолжают обитать в экстремальных условиях, таких как горячие источники и глубоководные гидротермальные источники. Это и есть «симбиогенез» — создание или генезис нового типа сущности из новой «симбиозы», или взаимовыгодных отношений, между уже существующими сущностями.
В такие моменты дерево жизни не просто разветвляется; оно также запутывается само в себе, его ветви сливаются, создавая радикально новые формы. Маргулис была одним из первых сторонников идеи о том, что эти события движут эволюционными скачками вперед.
Вероятно, что бактерии сами являются продуктом таких симбиотических событий — например, между РНК и метаболизмом. РНК могла бы размножаться без помощи белков, а метаболический мотор мог бы развиваться без помощи генов, но когда эти системы сотрудничают, они добиваются лучших результатов. Циклические сети химических реакций в тех черных курильщиках можно рассматривать как такое сотрудничество, набор реакций, которые, катализируя друг друга, могут образовать более устойчивое, самоподдерживающее целое.
Таким образом, в определенном смысле Дарвин мог быть прав, утверждая, что «это просто ерунда» — думать о происхождении жизни, поскольку жизнь могла не иметь единого начала, а скорее сплести себя из множества отдельных нитей, самые древние из которых выглядят как обычная химия. Умный замысел не требуется для того, чтобы это плетение происходило; достаточно неоспоримой логики, что иногда альянс создает что-то долговечное, и что все, что долговечно... остается.
Часто долговечность означает как создание, так и занятие совершенно новых ниш. Поэтому эукариоты не заменили бактерии; на самом деле, они в конечном итоге создали для них множество новых ниш. Точно так же симбиотическое возникновение многоклеточной жизни — еще один крупный эволюционный переход — не вытеснило одноклеточную жизнь.
Как древний пергамент, переписанный поколениями писцов, наша планета является палимпсестом, многослойное прошлое которого все еще различимо в настоящем. Даже черные курильщики, реликты ада, существовавшие миллиарды лет назад, продолжают бурлить на дне океана. Самокатализирующаяся химия протожизни, возможно, все еще варится там в глубинах, медленно, на океанском дне.
Идея о том, что эволюция движется за счет симбиотических слияний, самые ранние из которых предшествовали биологии в том виде, в каком мы ее знаем, имеет далеко идущие последствия. Одно из них заключается в том, что граница между жизнью и неживой природой нечетка; симбиогенез может включать любой процесс, который так или иначе самоподдерживается. Эволюционные динамики, таким образом, больше напоминают физический закон, чем биологический принцип.
Все подвержено эволюции, независимо от того, считаем ли мы это «живым» или нет.
Симбиогенетическая точка зрения также делает идею о четко определенных видах, классифицируемых по линнеевской таксономии, несколько неясной — или, в лучшем случае, ограниченной в применении. Такие таксономии предполагают, что происходит только разветвление, а не слияние. Бактерии, которые без стеснения передают гены даже между «видами», уже ставят под сомнение эту идею.
Таксономии полностью теряют смысл, когда мы пытаемся применить их к еще более текучим ситуациям, таким как возможная протожизнь в черных курильщиках, или микробиомы, или более сложные симбиозы, которые мы рассмотрим позже.

Финчевые с Галапагосов, из книги Дарвина,
«Путешествие на Бигле», 1845

Финчевые с Галапагосов, из книги Дарвина,
«Путешествие на Бигле», 1845

Финчевые с Галапагосов, из книги Дарвина,
«Путешествие на Бигле», 1845

Финчевые с Галапагосов, из книги Дарвина,
«Путешествие на Бигле», 1845
Возможно, самое важное, что симбиогенез объясняет стрелу времени эволюции, чего одна лишь классическая дарвиновская теория сделать не может.
Когда жизнь ветвится, специализируясь для адаптации к определенной нише — как у дарвиновских воробьиных с их различными формами клюва, каждая из которых оптимизирована для конкретного источника пищи — эти ветви, как правило, не более сложны, чем их предковая форма. Это привело некоторых классических эволюционных теоретиков к утверждению, что возрастающая сложность жизни на Земле является антропоцентрической иллюзией, ничем иным, как результатом случайного блуждания по генетическим возможностям.
Связано с этим, иногда можно услышать утверждение, что поскольку все существующие виды имеют равный возраст — как кажется, ведь все они происходят от общего одноклеточного предка около четырех миллиардов лет назад — ни один вид не является более «эволюционировавшим», чем другой.
На первый взгляд, это кажется разумным. Но, как мы уже видели, классическая дарвиновская теория испытывает трудности с объяснением того, почему жизнь, кажется, становится все более сложной, или, действительно, как жизнь могла возникнуть в первую очередь. Сами по себе случайные изменения или мутации могут лишь тонко настраивать, разнообразить и оптимизировать, позволяя проявление той или иной вариации, уже скрытой в генетическом пространстве. (Например, пространство возможных форм клюва воробьиных.)
Однако когда один прокариот оказывается внутри другого, или несколько клеток объединяются, чтобы создать многоклеточную жизнь, получившийся составной организм явно более сложен, чем его части. Возникает нечто поистине новое. Ветвление и тонкая настройка классической эволюции теперь могут начать действовать на совершенно другом уровне, в новом пространстве комбинаторных возможностей.
Классическая эволюция не ошибочна; она просто упускает половину истории — более быструю, более креативную половину. Можно сказать, что другой половиной эволюции является революция, и что революции происходят через симбиоз.
Знаково, что то же самое справедливо и для технологий. В палеолитические времена копье с рукояткой было не просто специализированной каменной точкой, но и чем-то новым, возникшим в результате сочетания как минимум трех частей: каменной точки, древка и чего-то вроде жилы, чтобы их связать.
Это также открыло новое пространство для комбинаторного дизайна. Или, в более современных терминах, кремниевый чип — это не просто эволюционировавший транзистор; это нечто новое, что можно создать, объединив несколько транзисторов на одном кристалле. Дизайн чипа не возникает просто из игры с параметрами дизайна отдельного транзистора.
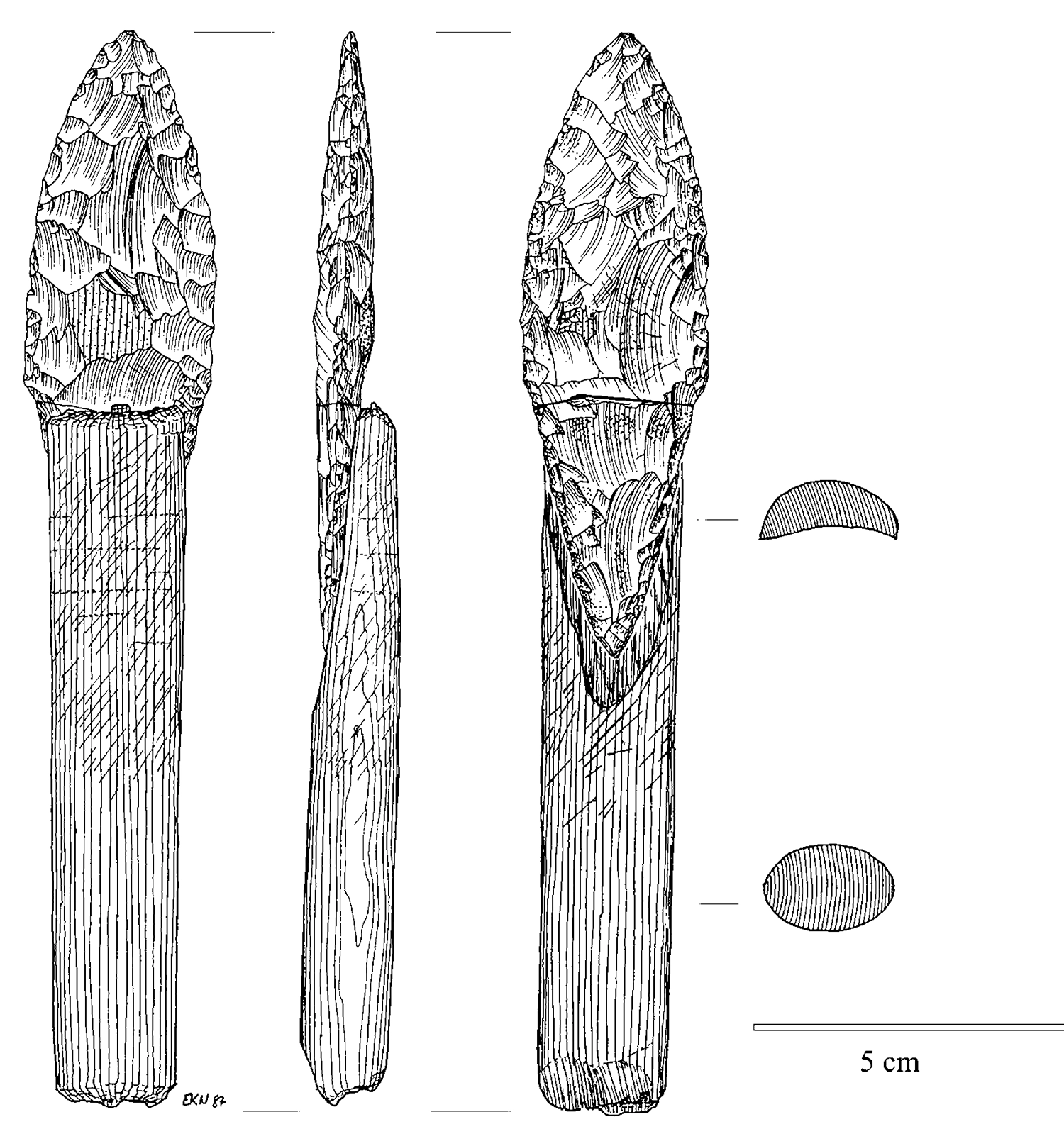 Наконечник копья с рукояткой из среднеголоценового арктического маленького инструмента с сайта Qeqertasussuk, Западная Гренландия. Нарисовано Эвой Кох, воспроизведено с разрешения Бьярне Грённова.
Вот почему эволюция движется от более простых форм к более сложным. Именно поэтому более простые формы часто продолжают существовать даже после успешного симбиогенетического события. Отдельные каменные наконечники, такие как ножи и ручные топоры, продолжали изготавливаться после изобретения копья с рукояткой; и, конечно, сами наконечники копий по-прежнему считаются каменными наконечниками.
Более того, независимо от того, насколько недавно был изготовлен тот или иной каменный наконечник, мы можем с уверенностью говорить о том, что каменные наконечники как категория «старше» копий. Копья должны были возникнуть позже, поскольку их существование зависело от предшествующего существования каменных наконечников.
Не менее значимо говорить о древних формах жизни, таких как бактерии и археи, сосуществующих наряду с более современными и гораздо более сложными, такими как люди, — при этом признавая, что люди, в определенном смысле, являются сложными колониями бактерий и архей, которые прошли через каскад симбиотических слияний.
Функции размножения
Хотя большинство биохимиков сосредоточили свои усилия на понимании конкретной истории и механизмов жизни на Земле, более общее понимание жизни пришло из неожиданной области: информатики. Теоретические основы этой удивительной связи восходят к двум основополагающим фигурам этой области — Алану Тьюрингу и Джону фон Нейману.
Наконечник копья с рукояткой из среднеголоценового арктического маленького инструмента с сайта Qeqertasussuk, Западная Гренландия. Нарисовано Эвой Кох, воспроизведено с разрешения Бьярне Грённова.
Вот почему эволюция движется от более простых форм к более сложным. Именно поэтому более простые формы часто продолжают существовать даже после успешного симбиогенетического события. Отдельные каменные наконечники, такие как ножи и ручные топоры, продолжали изготавливаться после изобретения копья с рукояткой; и, конечно, сами наконечники копий по-прежнему считаются каменными наконечниками.
Более того, независимо от того, насколько недавно был изготовлен тот или иной каменный наконечник, мы можем с уверенностью говорить о том, что каменные наконечники как категория «старше» копий. Копья должны были возникнуть позже, поскольку их существование зависело от предшествующего существования каменных наконечников.
Не менее значимо говорить о древних формах жизни, таких как бактерии и археи, сосуществующих наряду с более современными и гораздо более сложными, такими как люди, — при этом признавая, что люди, в определенном смысле, являются сложными колониями бактерий и архей, которые прошли через каскад симбиотических слияний.
Функции размножения
Хотя большинство биохимиков сосредоточили свои усилия на понимании конкретной истории и механизмов жизни на Земле, более общее понимание жизни пришло из неожиданной области: информатики. Теоретические основы этой удивительной связи восходят к двум основополагающим фигурам этой области — Алану Тьюрингу и Джону фон Нейману.
Алан Тьюринг в Принстонском университете
Я уже упоминал о работе Тьюринга 1950 года, в которой он представил Игра Подражания или Тест Тьюринга, но его первое великое достижение произошло пятнадцатью годами ранее. Получив степень бакалавра математики в Кембридже в 1935 году, Тьюринг сосредоточился на одной из ключевых нерешенных задач того времени: проблеме принятия решения (по-немецки Entscheidungsproblem), которая ставила вопрос, существует ли алгоритм для определения истинности произвольного математического утверждения.
Ответ оказался отрицательным, но способ, которым Тьюринг подошел к доказательству, оказался гораздо важнее самого результата — что было к счастью, поскольку американский математик Алонзо Чёрч уже опередил Тьюринга, опубликовав свое собственное доказательство несколько месяцев назад, используя другой подход.
После переписки с ним по поводу проблемы принятия решения, Тьюринг в конечном итоге переехал в Принстон, чтобы стать аспирантом Чёрча.
Доказательство математических утверждений о математических утверждениях требует определения общей нотации для таких утверждений и процедуры их оценки. Чёрч сделал это, используя математические символы, особенно строчную греческую букву λ (лямбда), тем самым изобретя самоссылающийся «лямбда-исчисление».
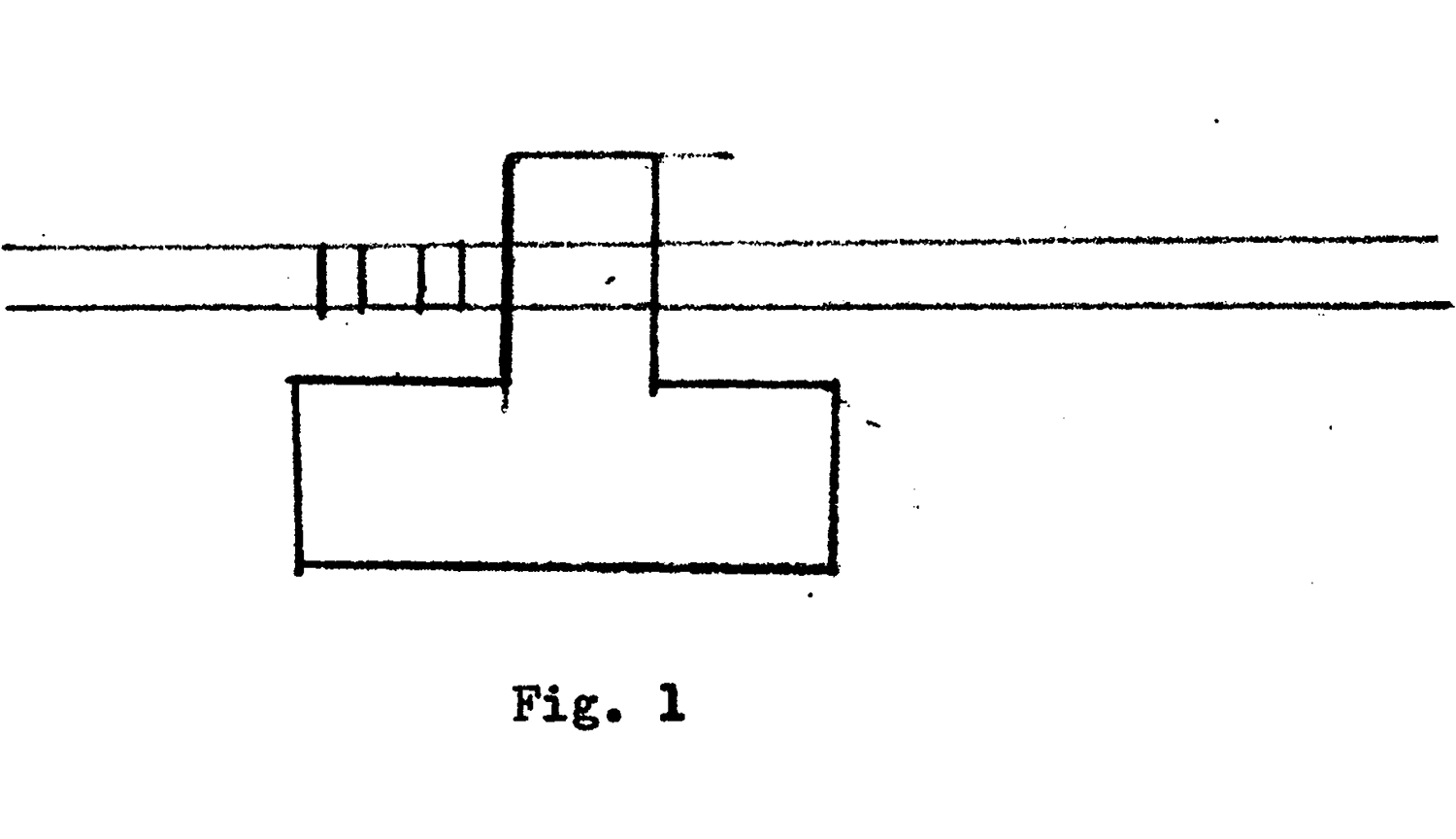
Вон Нейман иллюстрирует машину Тьюринга в начале своей третьей лекции 1945 года о высокоскоростных вычислительных машинах.
Тьюринг выбрал менее традиционный путь. Его процедура включала воображаемое устройство, которое мы теперь называем «машиной Тьюринга». Машина Тьюринга состоит из головки чтения/записи, которая может двигаться влево или вправо по бесконечной ленте, считывая и записывая символы на ленте в соответствии с набором правил, заданных встроенной таблицей.
Сначала Тьюринг показал, что такая машина может выполнять любые вычисления, которые можно сделать вручную, при условии наличия подходящей таблицы правил, достаточного времени и достаточной длины ленты.
Он предложил обозначение для записи таблицы правил, которую он назвал «Стандартным описанием» машины или «С. О.». Сегодня мы бы назвали это «программой».
А вот и действительно умная часть: если сама программа записана на ленте, существуют специальные таблицы правил, которые будут считывать программу и выполнять любые вычисления, которые она задает. Сегодня их называют «Универсальными машинами Тьюринга». В более точной формулировке Тьюринга: «Можно изобрести одну единственную машину, которая может использоваться для вычисления любой вычислимой последовательности. Если этой машине U предоставить ленту, на начале которой записано С. О. какой-либо вычислительной машины M, то U будет вычислять ту же последовательность, что и M».

Фотография удостоверения личности Джона фон Неймана, Лос-Аламос, 1940-е годы
Некоторые программы выполняются в течение определенного времени, а затем завершаются. Другие работают вечно. Поскольку Универсальная машина Тьюринга позволяет выражать сами программы как значения на ленте, программа может принимать другую программу в качестве входных данных. Можно представить себе такую «мета-программу», которая выводит один бит информации, указывающий, завершится ли введенная программа в конечном итоге или будет работать бесконечно. Тьюринг назвал это «проблемой остановки». В своей статье 1936 года он доказал, что программа, решающая проблему остановки для всех возможных входных программ — и которая сама гарантированно завершится — является логической невозможностью. Это был результат, который отрицательно ответил на
проблему решения.
Однако более значительным является то, что Универсальная машина Тьюринга определила универсальную процедуру для вычислений. Тьюринг вскоре смог доказать, что любая функция, которую можно вычислить с помощью его воображаемой машины, также может быть выражена в лямбда-исчислении Чёрча и наоборот — краеугольный камень идеи, ныне известной как тезис Чёрча-Тьюринга: вычислимость является универсальной концепцией, независимо от того, как она реализована.
В начале 1940-х годов Джон фон Нейман, венгерско-американский полимат, который уже сделал значительные вклады в физику и математику, обратил свое внимание на вычисления. Он стал ключевой фигурой в проектировании ENIAC и EDVAC — одних из первых в мире реальных универсальных машин Тьюринга, которые теперь просто называются «компьютерами».
Преобразование воображаемой математической машины в работающую физическую требовало множества концептуальных скачков, помимо значительных усилий в инженерии. Например, на протяжении многих лет было проявлено много креативности в поисках того, насколько простой может быть «Стандартное описание» универсальной машины Тьюринга. Для этого требуется всего несколько инструкций. Эзотерические языковые энтузиасты даже нашли способ вычислять с помощью единственной инструкции (так называемый OISC или «компьютер с одним набором инструкций»).
Тем не менее, существуют неразрешимые требования: инструкция или инструкции должны изменять окружение таким образом, чтобы последующие инструкции могли это «увидеть», и должно быть «условное ветвление», что означает, что в зависимости от состояния окружения произойдет либо одно, либо другое. В большинстве языков программирования это выражается с помощью операторов «если/то». Когда имеется только одна инструкция, она должна выполнять обе функции, как в языке SUBLEQ, единственной инструкцией которого является «вычесть и перейти, если результат меньше или равен нулю».

Программисты перед компьютером MANIAC, Лос-Аламосская национальная лаборатория, 1952 год
И Тьюринг, и фон Нейман были хорошо осведомлены о параллелях между компьютерами и мозгами. Отчет фон Неймана о EDVAC явно описывал основные строительные блоки машины, ее «логические элементы», как электронные нейроны.
Хотя эта аналогия и не выдерживает критики (как мы увидим, она не верна; нейроны гораздо сложнее логических элементов), его ключевое понимание заключалось в том, что как мозг, так и компьютеры определяются не своими механизмами, а тем, что они делают — их функцией, как в обыденном, так и в математическом смысле.
Однако в реальной жизни мозг не является абстрактной машиной, а частью тела, а тело — частью физического мира. Как можно говорить о функции живого организма исключительно в вычислительных терминах, когда он должен физически расти и размножаться?
Хотя они работали независимо друг от друга, к концу своей жизни Тьюринг и фон Нейман оба были очарованы связью между биологией и вычислениями. Тьюринг провел пионерские исследования в области «морфогенеза», выясняя, как клетки могут использовать химические сигналы, которые он назвал «морфогенами», для формирования сложных самоорганизующихся паттернов — ключа к многоклеточности. Хотя компьютеры все еще были слишком примитивными, чтобы выполнять детализированные симуляции таких систем, он математически показал, как так называемые «реакционно-диффузионные» уравнения могут генерировать пятна, подобные тем, что есть у леопардов и коров, или управлять ростом щупалец, как у пресноводного полипа, гидры.

Паттерн реакционно-диффузионного процесса из работы Тьюринга «Химическая основа морфогенеза», 1952
Примерно в то же время, в статье 1951 года, фон Нейман представил себе машину, состоящую из стандартных частей, как детали конструктора Lego, которая плавала по резервуару, где эти части могли бы находиться на поверхности воды. Задачей машины было собрать все необходимые детали и построить другую машину, подобную себе. Конечно, именно это делает бактерия для размножения; на самом деле, это то, что каждая клетка должна делать, чтобы делиться, и что каждая мать должна делать, чтобы родить.

Записка о самовоспроизводящихся машинах из записной книжки пионера кибернетики У.
Росс Эшби, 22 декабря 1954 года
Простая структура, как кристаллическое семя, может «воспроизводиться», просто служа шаблоном для кристаллизующегося вокруг нее материала. Но сложная машина — например, имеющая внутренние компоненты — не может быть своим собственным шаблоном. И если вы являетесь сложной машиной, то, на первый взгляд, создание чего-то столь же сложного, как и вы сами, кажется парадоксом, подобным попытке поднять себя за собственные шнурки. Тем не менее, фон Нейман показал, что это не только возможно, но и вполне осуществимо, используя обобщение универсальной машины Тьюринга.
Он представил «машину А», которая будет считывать ленту с последовательными инструкциями по сборке, основанными на ограниченном каталоге деталей, и выполнять их шаг за шагом. Затем «машина Б» скопирует ленту — при условии, что сама лента также состоит из доступных деталей. Если инструкции по созданию машин А и Б закодированы на ленте, то вуаля — вы получите репликатор.
Инструкции для создания любых дополнительных не репродуктивных машин также могут быть закодированы на ленте, так что репликатор сможет создать что-то более сложное, чем он сам.
Семя или оплодотворенная яйцеклетка иллюстрируют эту идею. Более того, кодирование инструкций для самосоздания в форме, которая сама воспроизводится (лента), является ключом к открытой эволюционной способности, что означает возможность эволюции выбирать произвольные изменения в дизайне, которые будут наследоваться следующему поколению.
Удивительно, но фон Нейман описал эти требования для эволюционной, самовоспроизводящейся машины еще до открытия структуры и функции ДНК. Тем не менее, он все понял совершенно верно. Для жизни на Земле ДНК — это лента; ДНК-полимераза, которая копирует ДНК, — это «машина Б»; а рибосомы, которые создают белки, следуя последовательным инструкциям, закодированным в ДНК, — это «машина А».
Рибосомы и ДНК-полимераза состоят из белков, последовательности которых, в свою очередь, закодированы в нашей ДНК и производятся рибосомами. Так жизнь поднимает себя за собственные «петли».
Жизнь как вычисление
Хотя это редко полностью осознается, интуиция фон Неймана установила глубокую связь между жизнью и вычислениями. Помните, машины A и B — это машины Тьюринга. Они должны выполнять инструкции, которые влияют на их окружение, и эти инструкции должны выполняться в цикле, начиная с начала и заканчивая в конце. Это требует разветвлений, таких как: «если следующая инструкция — кодон CGA, тогда добавь аргинин к строящемуся белку» и «если следующая инструкция — UAG, тогда СТОП». Называть ДНК «программой» — это не метафора, это буквально так.
Конечно, существуют значительные различия между биологическими вычислениями и теми цифровыми вычислениями, которые выполняет ENIAC или ваш смартфон. ДНК тонка и многослойна, включая такие явления, как эпигенетика и эффекты близости генов. Клеточная ДНК — это далеко не вся история. Наши тела содержат (и постоянно обмениваются) бесчисленными бактериями и вирусами, каждый из которых выполняет свой собственный код.
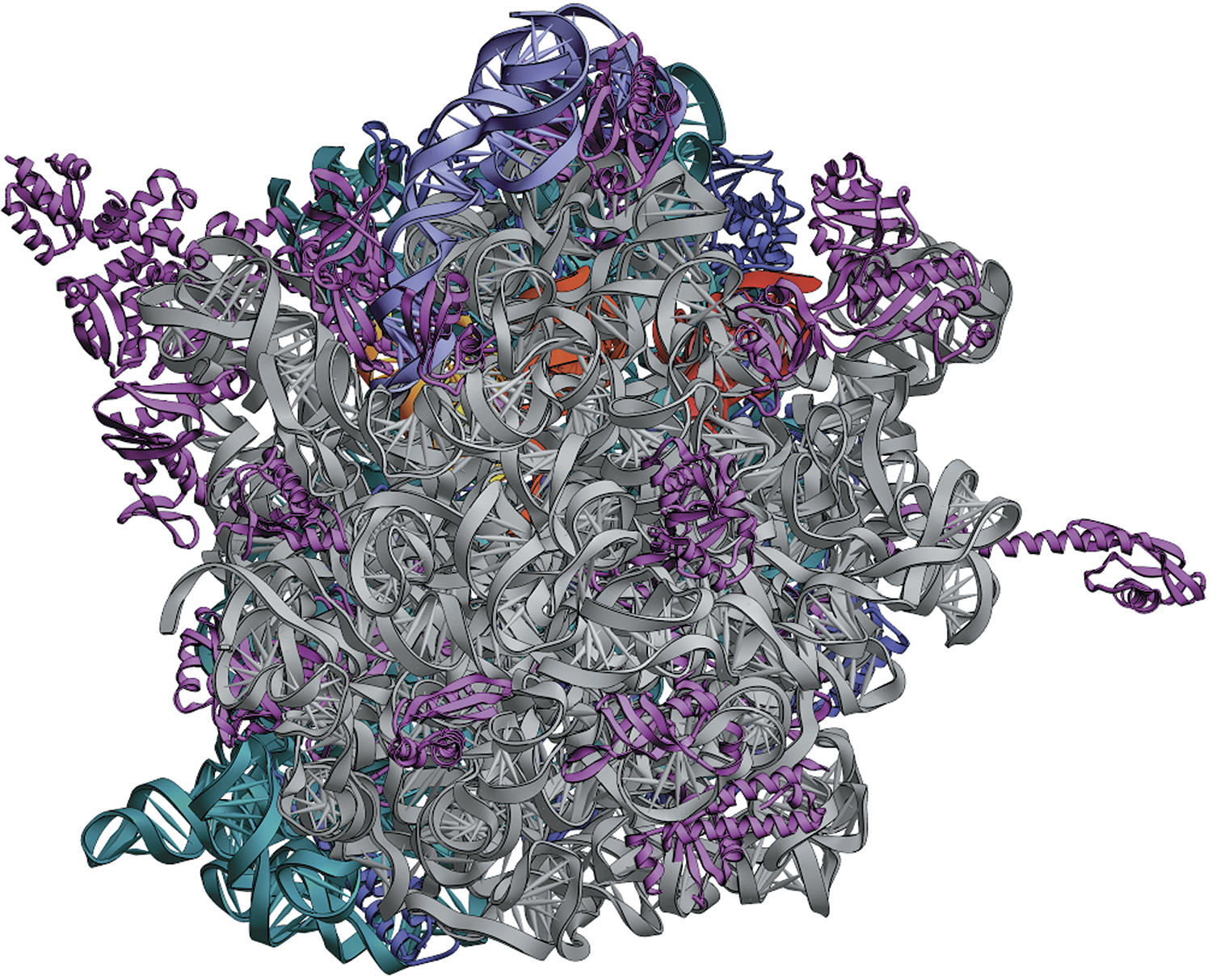
Структура рибосомы 70S
Биологические вычисления «масштабно параллельны», децентрализованы и шумны. В ваших клетках находится около трехсот квинтильонов рибосом, работающих одновременно. Каждая из этих изысканно сложных плавающих фабрик белков является, по сути, крошечным компьютером — хотя и стохастическим, что означает, что его поведение не совсем предсказуемо. Движения шарнирных компонентов, захват и высвобождение меньших молекул, а также манипуляции с химическими связями все индивидуально случайны, обратимы и неточны, подвержены постоянным тепловым колебаниям.
Только статистическая асимметрия позволяет одной стороне преобладать над другой, при этом хитрые движения оригами склонны «закреплять» определенные шаги, так что следующий шаг становится вероятным. Это сильно отличается от работы логических вентилей в компьютере, которые являются необратимыми и разработаны с надежностью и воспроизводимостью на уровне девяноста девяти с лишним процентов. Тем не менее, биологические вычисления — это тоже вычисления. И использование случайности здесь является особенностью, а не ошибкой. На самом деле многие классические алгоритмы в информатике также требуют случайности (хотя и по другим причинам), что может объяснить, почему Тьюринг настаивал на том, чтобы Ferranti Mark I, ранний компьютер, который он помог спроектировать в 1951 году, включал инструкцию для генерации случайных чисел. Таким образом, случайность представляет собой небольшое, но важное концептуальное расширение оригинальной машины Тьюринга, хотя любой компьютер может смоделировать это, вычисляя детерминированные, но выглядящие случайными или «псевдослучайными» числа.

Тьюринг (справа) за консолью компьютера Ferranti Mark II
Параллелизм также становится все более фундаментальным для вычислений сегодня. Современный ИИ, например, зависит как от массового параллелизма, так и от случайности — как в параллелизованном алгоритме «стохастического градиентного спуска» (SGD), который используется для обучения большинства современных нейронных сетей, так и в настройке «температуры», применяемой в чат-ботах для введения степени случайности в их вывод, а также в параллелизме графических процессоров (GPU), которые обеспечивают работу большинства ИИ в дата-центрах.
Традиционные цифровые вычисления, которые полагаются на централизованное, последовательное выполнение инструкций, были продуктом технологических ограничений. Первые компьютеры должны были выполнять длинные вычисления, используя как можно меньше деталей. Изначально эти детали представляли собой ненадежные, дорогие вакуумные лампы, которые имели тенденцию выходить из строя и требовали частой замены вручную. Естественным дизайном, таким образом, стал минимальный «Центральный процессор» (ЦП), работающий с последовательностями битов, которые перемещались туда и обратно из внешней памяти.
Это стало известно как «архитектура фон Неймана».
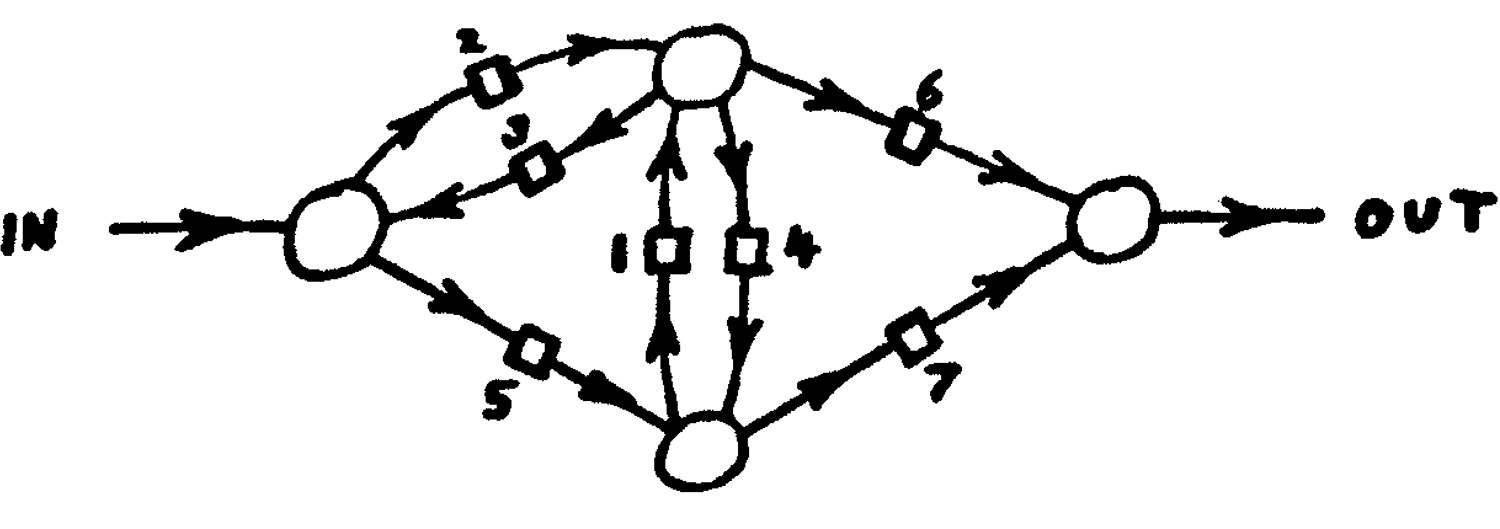
Иллюстрация Тьюринга «неорганизованной машины» (или нейронной сети) из «Интеллектуальной машины», 1948 год.
Тем не менее, Тьюринг и фон Нейман оба осознавали, что вычисления могут осуществляться и другими способами. Модель морфогенеза Тьюринга была биологически вдохновленной формой массово параллельных, распределенных вычислений. То же самое касалось и его ранней концепции «неорганизованной машины» — случайно соединенной нейронной сети, смоделированной по образцу мозга младенца.
Это были видения того, как могли бы выглядеть вычисления без центрального процессора — и как они действительно выглядят в живых системах.
Фон Нейман также начал исследовать массово параллельные подходы к вычислениям еще в 1940-х годах. В обсуждениях с польским математиком Станиславом Уламом в Лос-Аламосе он задумал идею «клеточных автоматов» — сеток, состоящих из простых вычислительных единиц, каждая из которых подчиняется одному и тому же правилу и одновременно изменяет свои состояния, общаясь только с ближайшими соседями. С характерной смелостью фон Нейман даже разработал на бумаге ключевые компоненты самовоспроизводящегося клеточного автомата, включая горизонтальную линию клеток, представляющую собой «ленту», и блоки клеточной «схемы», реализующие машины A и B.
Универсальный конструктор фон Неймана из
«Теории самовоспроизводящихся автоматов», 1967 год.
Проектирование клеточного автомата гораздо сложнее, чем обычное программирование, поскольку каждая клетка или «пиксель» одновременно изменяет свое состояние и окружающую среду. Когда такой параллелизм работает на многих масштабах одновременно и сочетается со случайностью и тонкими эффектами обратной связи, как в биологических вычислениях, становится еще труднее рассуждать о нем, «программировать» или «отлаживать».
Тем не менее, мы должны помнить, что эти два пионера очень четко поняли: вычисления не обязательно должны выполняться с помощью центрального процессора, логических вентилей, двоичной арифметики или последовательных программ. Существует бесконечное множество способов вычисления. Тьюринг и его последователи показали, что все они эквивалентны, что является одним из величайших достижений теоретической информатики.
Это «независимость платформы» или «многообразие реализаций» означает, что любой компьютер может эмулировать любой другой. Однако если компьютеры имеют разные конструкции, эмуляция может происходить медленно. Например, самовоспроизводящийся клеточный автомат фон Неймана никогда не был физически построен — хотя было бы интересно это увидеть! Впервые его эмулировали только в 1994 году, почти через полвека после его проектирования.

Реализация самовоспроизводящегося клеточного автомата фон Неймана, Пезавенто, 1995. Второе поколение почти завершило строительство третьего (линии, уходящие вправо, — это инструкции, значительно длиннее, чем показано).
Это не могло произойти намного раньше. Последовательному компьютеру требуется серьезная вычислительная мощность, чтобы пройти через 6,329 клеток автомата за шестьдесят три миллиарда временных шагов, необходимых для завершения его репродуктивного цикла. На экране он работал как и ожидалось: пиксельная двумерная машина Руба Голдберга, сидящая на 145,315 клеточной инструкции, уходящей вправо, выкачивающая информацию из ленты и протягивающая «письменную руку», чтобы медленно напечатать работающий клон себя чуть выше и справа от оригинала.
Аналогично, для последовательного компьютера неэффективно эмулировать параллельную нейронную сеть, наследницу «неорганизованной машины» Тьюринга. В результате запуск больших нейронных сетей, таких как те, что используются в чат-ботах на основе трансформеров, стал практичным лишь недавно благодаря постоянному прогрессу в миниатюризации, скорости и параллелизме цифровых компьютеров.
В 2020 году мой коллега Алекс Мордвинцев разработал умное сочетание современных нейронных сетей, морфогенеза Тьюринга и клеточных автоматов фон Неймана. Создание Алекса, «нейронный клеточный автомат» (NCA), заменяет простое правило обработки пикселей классического клеточного автомата на нейронную сеть. Эта сеть, способная воспринимать и изменять несколько значений, представляющих локальные концентрации морфогенов, может быть обучена «выращивать» любой желаемый узор или изображение, а не только зебровые полосы или пятна ягуара.
Нейронный клеточный автомат, генерирующий (или регенерирующий) эмодзи ящерицы, из работы Мордвинцева и др. 2020 года.
Настоящие клетки не содержат нейронных сетей в буквальном смысле, но они выполняют высокоразвитые, нелинейные и целенаправленные «программы», чтобы решать, какие действия предпринять в ответ на внешние стимулы и внутреннее состояние. NCA предлагает общий способ моделирования диапазона возможного поведения клеток, чьи действия не связаны с движением, а только с изменениями состояния (в данном случае, представленными цветом) и поглощением или выделением химических веществ.
Первый NCA, который Алекс показал мне, изображал эмодзи ящерицы, способной регенерировать не только свой хвост, но и конечности и голову! Это была мощная демонстрация того, как сложная многоклеточная жизнь может «думать локально», но «действовать глобально», даже когда каждая клетка (или пиксель) выполняет одну и ту же программу — так же, как каждая из ваших клеток использует одну и ту же ДНК. Это было наше первое погружение в область, известную сегодня как «искусственная жизнь» или «ALife».
Искусственная жизнь
Работы фон Неймана о самовоспроизводящихся автоматах показывают нам, что в вселенной, физические законы которой не допускают вычислений, жизнь не могла бы эволюционировать. К счастью, физика нашей вселенной позволяет вычисления, что подтверждается тем фактом, что мы можем строить компьютеры — и что мы вообще здесь находимся. Теперь мы можем задать вопрос: в вселенной, способной к вычислениям, как часто будет возникать жизнь? Очевидно, что это произошло здесь.
Чудо, неизбежность или что-то среднее?
Несколько моих коллег и я решили исследовать этот вопрос в конце 2023 года.
Наши первые эксперименты проводились с использованием эзотерического языка программирования, который был изобретен тридцать лет назад швейцарским студентом-физиком и любителем жонглирования Урбаном Мюллером. Боюсь, он назвал этот язык... Brainfuck. Все комментарии по поводу названия направляйте к нему.
Тем не менее, название полностью соответствует сути; программировать на этом языке — настоящее испытание. Вот, например, программа на Brainfuck, которая выводит «helloworld» — и удачи вам с её пониманием:
++++++[−>+++++<]>−[>[++++>]++++[<]>−]>>>>.>+.<<…<−.<+++.>.+++.>.>>−.
Плюс Brainfuck заключается в его абсолютном минимализме. Это не совсем язык с единственной инструкцией, как SUBLEQ, но, как вы видите, он включает лишь несколько операций. Подобно машине Тьюринга, он задает головку для чтения/записи, которая может перемещаться влево (инструкция «<») или вправо (инструкция «>») по ленте. Инструкции «+» и «−» увеличивают и уменьшают значение байта в текущей позиции на ленте.
Инструкции «,» и «.» вводят байт с консоли или выводят байт на консоль (в приведенном выше коде вы можете посчитать десять инструкций «.», по одной для каждой буквы в «helloworld»). Наконец, инструкции «[» и «]» реализуют циклы: «[» пропустит вперед к соответствующей «]», если байт в текущей позиции равен нулю, а «]» вернется к соответствующей «[», если байт не равен нулю. Вот и все!
Трудно поверить, что Brainfuck может быть использован для полной реализации, скажем, операционной системы Windows, но он действительно является «полностью Тьюрингом». Это означает, что при достаточном времени и памяти (то есть при достаточно длинной ленте) он может эмулировать любую другую компьютерную систему и вычислить всё, что можно вычислить.
В нашей версии, которую мы называем bff, есть «суп», содержащий тысячи лент, каждая из которых включает как код, так и данные. Это ключевой момент: в «классическом» Brainfuck код отделен от ленты, тогда как в bff мы хотели, чтобы код мог модифицировать сам себя.
Это может произойти только в том случае, если код сам находится на ленте, как изначально задумал Тьюринг.
Ленты bff имеют фиксированную длину — 64 байта, примерно такую же, как загадочная программа «helloworld», упомянутая выше. Сначала они заполнены случайными байтами. Затем они взаимодействуют случайным образом, снова и снова. В ходе взаимодействия две случайно выбранные ленты соединяются, и эта объединенная лента длиной 128 байт запускается, потенциально изменяя саму себя. Затем 64-байтовые половины снова разделяются и возвращаются в «суп». Время от времени значение байта случайным образом изменяется, как это делает космическое излучение с ДНК.
Поскольку bff имеет всего семь инструкций (представленных символами «<>+−,[ ]») и существует 256 возможных значений байтов, после случайной инициализации только 7/256, или 2,7 процента, байтов на данной ленте будут содержать действительные инструкции; любые недействительные инструкции просто игнорируются.
Таким образом, в начале взаимодействия между лентами не дает много результатов. Время от времени действительная инструкция изменяет байт, и это изменение сохраняется в «супе». В среднем, однако, за одно взаимодействие происходит лишь пара вычислительных операций, и обычно они не оказывают никакого эффекта. Другими словами, хотя в этой игрушечной вселенной теоретически возможно любое вычисление, на практике происходит очень немного — поначалу. Случайная мутация может изменить байт здесь и там. Даже когда действительная инструкция вызывает изменение байта, это изменение произвольно и бессмысленно.
Но после миллионов взаимодействий происходит нечто волшебное: ленты начинают воспроизводиться! Когда они порождают копии самих себя и друг друга, случайность уступает место сложному порядку. Внезапность этого изменения напоминает «фазовый переход» в материи, как между газом и жидкостью или жидкостью и твердым телом. Действительно, начальный хаос «супа» очень похож на хаос случайно движущихся молекул газа.
Таким образом, случайный, нефункциональный код получил название «туринг-газ»; однако в bff он «конденсируется» в работающий код, что представляет собой нечто гораздо более сложное, чем твердое или жидкое.
▶ Ранний запуск bff, показывающий эволюцию от случайной инициализации до полной репликации на ленте; показаны только байты, кодирующие инструкции.
Мы могли бы назвать это «компьютронием», поскольку в момент фазового перехода объем вычислений стремительно возрастает — помните, воспроизводство требует вычислений. Две из семи инструкций Brainfuck («[» и «]») предназначены для условного ветвления и определяют циклы в коде. Воспроизводство требует как минимум одного такого цикла («копировать байты до завершения»), что приводит к тому, что количество выполняемых инструкций за одно взаимодействие достигает тысяч.
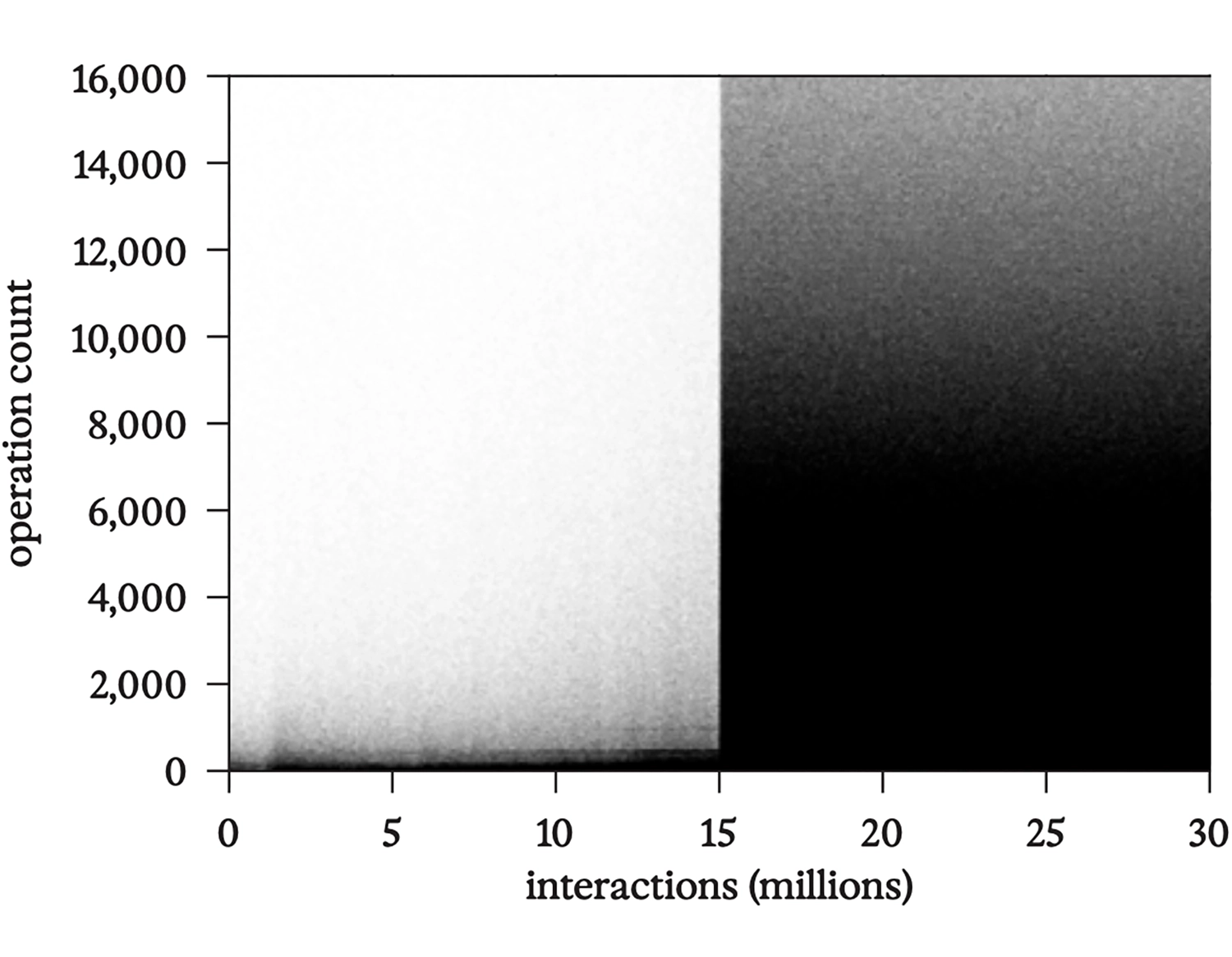
Диаграмма рассеяния, показывающая первые тридцать миллионов взаимодействий в запуске bff в виде точек, высота которых равна количеству операций. Обратите внимание на резкий переход к вычислительно интенсивной полной репликации на ленте при пятнадцати миллионах взаимодействий.
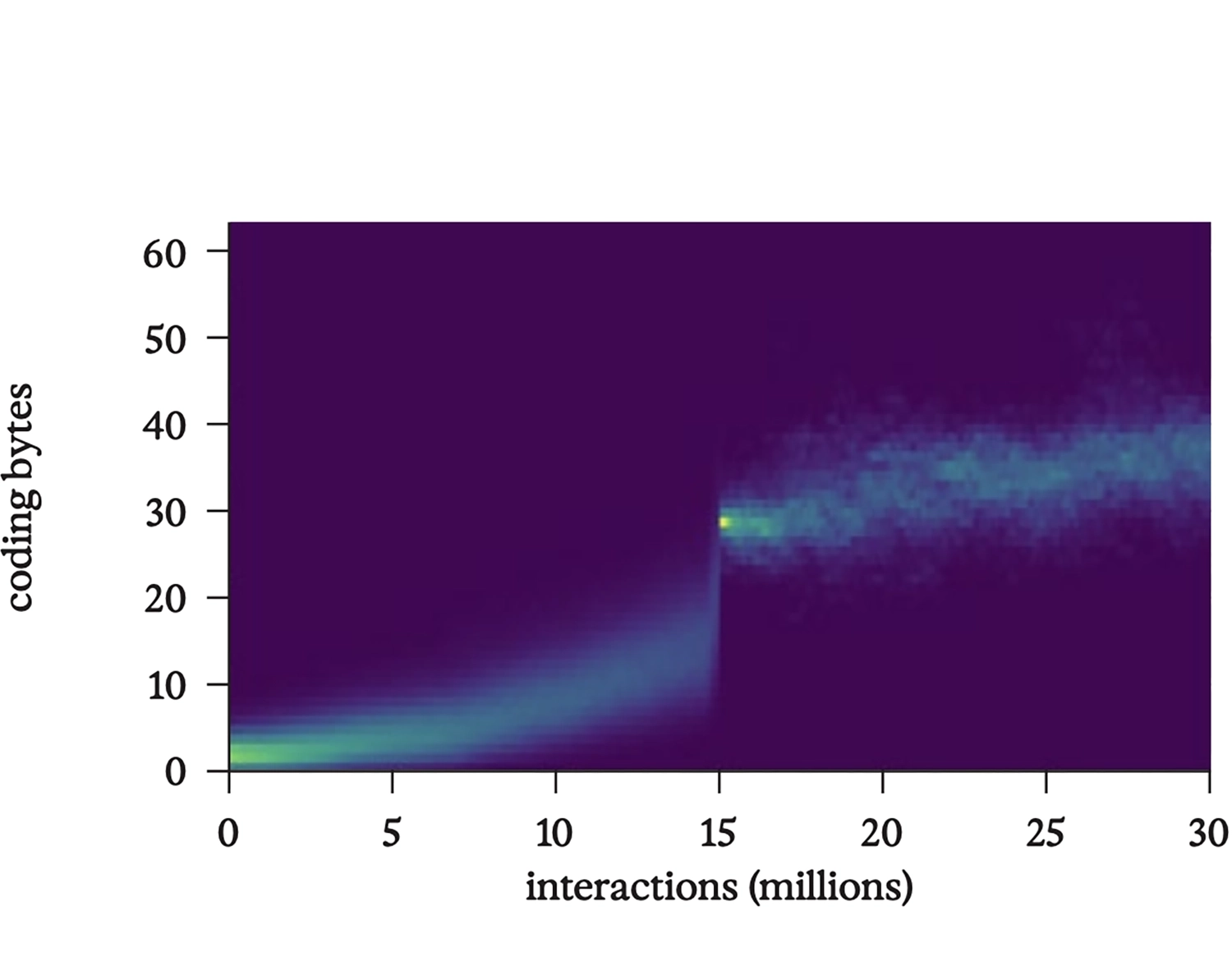
Гистограмма, показывающая количество байтов, кодирующих инструкции, на каждой ленте в том же запуске bff; обратите внимание на продолжающийся рост объема кода даже после перехода к полной репликации на ленте при пятнадцати миллионах взаимодействий.
Код больше не случайный, а явно целенаправленный, в том смысле, что его функцию можно анализировать и обратным образом разрабатывать. Неудачная мутация может его сломать, сделав неспособным к воспроизводству. Со временем код эволюционирует, вырабатывая умные стратегии для повышения своей устойчивости к таким повреждениям.
Это возникновение функции и цели похоже на то, что мы наблюдаем в органической жизни на всех уровнях; мы можем говорить о функции кровеносной системы, почки или митохондрии и о том, как они могут «отказать» — хотя никто не проектировал эти системы.
Мы воспроизвели наш основной результат с использованием различных языков программирования и сред.
Алекс (известный благодаря нейронным клеточным автоматам) создал еще один прекрасный микс, на этот раз между клеточными автоматами и bff. Каждый из 200×200 "пикселей" содержит программную ленту, а взаимодействия происходят только между соседними лентами на сетке. В знак уважения к нашим nerd-детствам, ленты интерпретируются как инструкции для культового микропроцессора Zilog Z80, выпущенного в 1976 году и использовавшегося во многих 8-битных компьютерах на протяжении лет (включая Sinclair ZX Spectrum, Osborne 1 и TRS-80). И здесь тоже из случайных взаимодействий вскоре появляются сложные репликаторы, эволюционируя и распространяясь по сетке в последовательных волнах.
▶ Эволюция конкурирующих репликаторов на сетке 200×200, смоделированных с использованием процессоров Zilog Z80
Наши симуляции показывают, что в целом жизнь возникает спонтанно, когда условия это позволяют. Эти условия кажутся минимальными: достаточно лишь среды, способной поддерживать вычисления, немного случайности и достаточно времени.
Давайте остановимся и подумаем, почему это так замечательно.
На интуитивном уровне не ожидаешь, что функция или целенаправленность возникнут спонтанно. Конечно, мы давно знаем, что скромная степень порядка может возникнуть из изначально случайных условий; например, набегание волн может примерно отсортировать песок на пляже, создавая градиент от мелкого к крупному. Но если бы мы начали с песка на пляже, подвергнутого случайному воздействию волн, и вернулись через несколько часов, чтобы обнаружить там написанное стихотворение или песчинки, слившиеся в сложную электронную схему, мы бы предположили, что кто-то с нами шутит.
Экстремальная маловероятность спонтанного возникновения сложного порядка обычно понимается как следствие термодинамики, раздела физики, занимающегося статистическим поведением материи, подверженной случайным тепловым колебаниям — то есть, всей материи, поскольку при температуре выше абсолютного нуля всё подвержено такой случайности. Материя, подвергающаяся случайным силам, должна становиться более случайной, а не менее.
Тем не менее, жизнь, растущая, размножающаяся, эволюционирующая и вообще существующая, кажется, нарушает этот принцип.
Однако это нарушение лишь на первый взгляд, ведь жизнь требует притока свободной энергии, что позволяет сдерживать силы энтропии. Тем не менее, кажущееся спонтанным возникновение и «комплексизация» живых систем, по всей видимости, не строго запрещены физическими законами, но, по крайней мере, не объясняются ими. Именно поэтому великий физик Эрвин Шрёдингер (1887–1961) написал в своем влиятельном небольшом труде, опубликованном в 1944 году под названием «Что такое жизнь?»:
«Живая материя, хотя и не уклоняется от “законов физики”, установленных на сегодняшний день, вероятно, включает в себя “другие законы физики”, ранее неизвестные, которые, однако, после их раскрытия станут такой же неотъемлемой частью этой науки, как и прежние».
Термодинамика
Прежде чем перейти к тем «другим законам физики», полезно более внимательно рассмотреть оригинальные законы, особенно Второй закон термодинамики.
Это глубокие воды. Хотя основные идеи восходят к революционной работе математического физика XIX века Людвига Больцмана (1844–1906), мы можем понять их суть без математики. Тем не менее, концептуально сложные идеи Больцмана ставили в тупик многих его коллег-ученых, и последствия его работы продолжают вызывать споры даже сегодня. Многое было сказано о том, как Эйнштейн перевернул наши повседневные представления о пространстве и времени с ног на голову с помощью своей теории относительности, разработанной в ее первоначальной форме в 1905 году — всего за год до того, как Больцман, страдая от биполярного расстройства, покончил с собой. Тем не менее, можно утверждать, что более ранние идеи Больцмана еще радикальнее подрывают наши интуиции о времени, причине и следствии, чем теория относительности Эйнштейна.
Давайте погрузимся в это.
Ржавые и изъеденные стержни частично обрушенного семидесятилетнего Железного моста через реку Нанду, провинция Хайнань, Китай
Второй закон термодинамики утверждает, что любая замкнутая система со временем будет увеличивать свою энтропию, становясь все более неупорядоченной. Например, ручная газонокосилка изначально представляет собой прекрасно отполированную машину с острыми спиральными лезвиями, круглыми колесами и зубчатыми шестернями, все это соединено на плавно вращающихся подшипниках. Если оставить ее на улице, подшипники закиснут, лезвия затупятся, и начнется окисление. Спустя некоторое время останется лишь груда ржавчины.
Аналогично, если вы возьмете мертвую бактерию (которая, хоть и гораздо меньше, но значительно сложнее, чем ручная газонокосилка) и опустите ее в колбу с водой, ее клеточная мембрана в конечном итоге разложится, различные части высыплются наружу, и через некоторое время останутся лишь простые молекулы, равномерно распределенные по колбе.
Второй закон придает времени направление, поскольку фундаментальные законы физики в нашей вселенной почти что обратимы по времени.
Странно, но правда: уравнения Ньютона (классическая динамика), уравнения Максвелла (электромагнетизм), уравнения Шрёдингера (квантовая физика), уравнения Эйнштейна (специальная и общая теория относительности) — все эти физические законы работали бы так же, если бы время шло в обратном направлении. Разница между прошлым и будущим исчезает, когда мы рассматриваем только математику динамических законов или отдельное микроскопическое событие во вселенной. Различие между прошлым и будущим, или причиной и следствием, становится заметным только тогда, когда мы отдаляемся от этого микроскопического мира отдельных событий и вместо этого рассматриваем их статистику.
Вот распространенный мыслительный эксперимент для студентов, чтобы проиллюстрировать эту точку зрения на примере ньютоновской динамики. Представьте себе видеозапись шаров на бильярдном столе, все они находятся в случайных позициях и движутся в случайных направлениях.
Они столкнутся друг с другом и с бортами стола, отскакивая под новыми углами. Если мы (хотя и нереалистично) предположим движение без трения и полностью упругие столкновения (т.е. шары не замедляются при катании, и никакая энергия столкновения не рассеивается в виде тепла), это будет продолжаться вечно. Суммарные импульсы и суммарные энергии шаров останутся постоянными — и будет невозможно определить, смотрите ли вы видео вперед или назад. В такой вселенной причинность теряет смысл, потому что ничто не отличает причины от следствий.
Если начальные условия были случайными, то позиции шаров будут продолжать случайным образом распределяться по поверхности стола, пока они все отскакивают друг от друга, передавая часть своей энергии при каждом столкновении. Астрономически маловероятно, что шары соберутся в одном месте. Их скорости также будут случайно распределены как по направлению, так и по величине, так что также будет астрономически маловероятно, что они, например, вдруг начнут двигаться точно параллельно краям стола. Полный беспорядок, другими словами, является стабильным равновесием.
При наличии любого теплового шума (т.е. случайных возмущений) это единственное стабильное равновесие.
Предположим, однако, что происходит почти невозможное. Вы видите, как пятнадцать шаров сходятся в идеальную, неподвижную треугольную решетку, каждый шар касается своих соседей, в то время как биток, поглотив всю комбинированную кинетическую энергию остальных пятнадцати, стремительно уносится от треугольника. Ах! Теперь мы знаем, что наблюдаем за началом игры в пул — и мы знаем, что смотрим на это в обратном порядке. Стрела времени установлена. Вместе с ней возникает причинность: треугольник разрушился потому, что в него врезался биток.
▶
Бильярдные шары в движении: вперед или назад во времени?
Теоретически ничто не мешает тому, чтобы точная последовательность столкновений привела к тому, что вся энергия передается одному шару, в то время как остальные остаются расположенными в идеальном треугольнике; но статистически вероятность того, что такое упорядоченное состояние возникнет из беспорядка, стремится к нулю —
если только, возможно, какой-то гений не запустил шары в движение именно так.
Хотя ключевые термодинамические концепции были разработаны только в девятнадцатом веке, вера эпохи Просвещения в Бога, который запустил вселенную именно так, интуитивно возникает из кажущейся невозможности порядка, возникающего спонтанно из беспорядка.
В механической вселенной бильярдного стола, где законы физики неприкосновенны, а законы статистики, кажется, неумолимо разрушают любой предшествующий порядок с течением времени, кажется абсурдным, что что-то столь сложное, как жизнь, могло бы возникнуть спонтанно без какого-либо сверхъестественного агента, действующего как «первопричина». Только Бог с изысканным предвидением мог «инициализировать» Большой Взрыв так, что в океанах Земли миллиарды лет назад простые органические молекулы, плавающие, казалось бы, наугад, могли соединиться в работающую бактерию —
невероятность, которая в миллионы раз меньше, чем вероятность того, что пятнадцать из шестнадцати летающих бильярдных шаров спонтанно образуют неподвижный треугольник.
Вселенная бильярдных шаров, которую я только что описал, может показаться абстрактной или произвольной, но теоретики девятнадцатого века, такие как Больцман, заинтересовались этой проблемой по самым практическим причинам: это была физика, стоящая за паровой энергией, а значит, и за всей Промышленной революцией.
Инженерия предшествовала теории, как это часто бывает.
Томас Ньюкомен (1664–1729), английский изобретатель и баптистский проповедник, разработал первый практический топливный двигатель в 1712 году. Он основывался на цикле нагрева и охлаждения.
Сначала пар из котла поступал в цилиндрическую камеру, поднимая поршень; затем клапан пара закрывался, и открывался второй клапан, впрыскивающий струю холодной воды, что вызывало конденсацию пара и опускало поршень обратно. Когда поршень поднимался и опускался, он раскачивал гигантскую балку взад-вперед, которая, в оригинальном проекте Ньюкомена, использовалась для откачки воды из затопленных шахт (которые, в свою очередь, обеспечивали углем эти двигатели, которые вскоре начали потреблять его с жадностью).
Шотландский изобретатель и предприниматель Джеймс Уатт (1736–1819) значительно улучшил конструкцию парового двигателя в 1776 году, сделав его практичной заменой человеческой и животной силы в широком спектре применений. Именно тогда началась настоящая Индустриальная революция; впервые человек-made машины начали метаболизировать в большом масштабе, «поедая» сложные органические молекулы для выполнения механической работы.
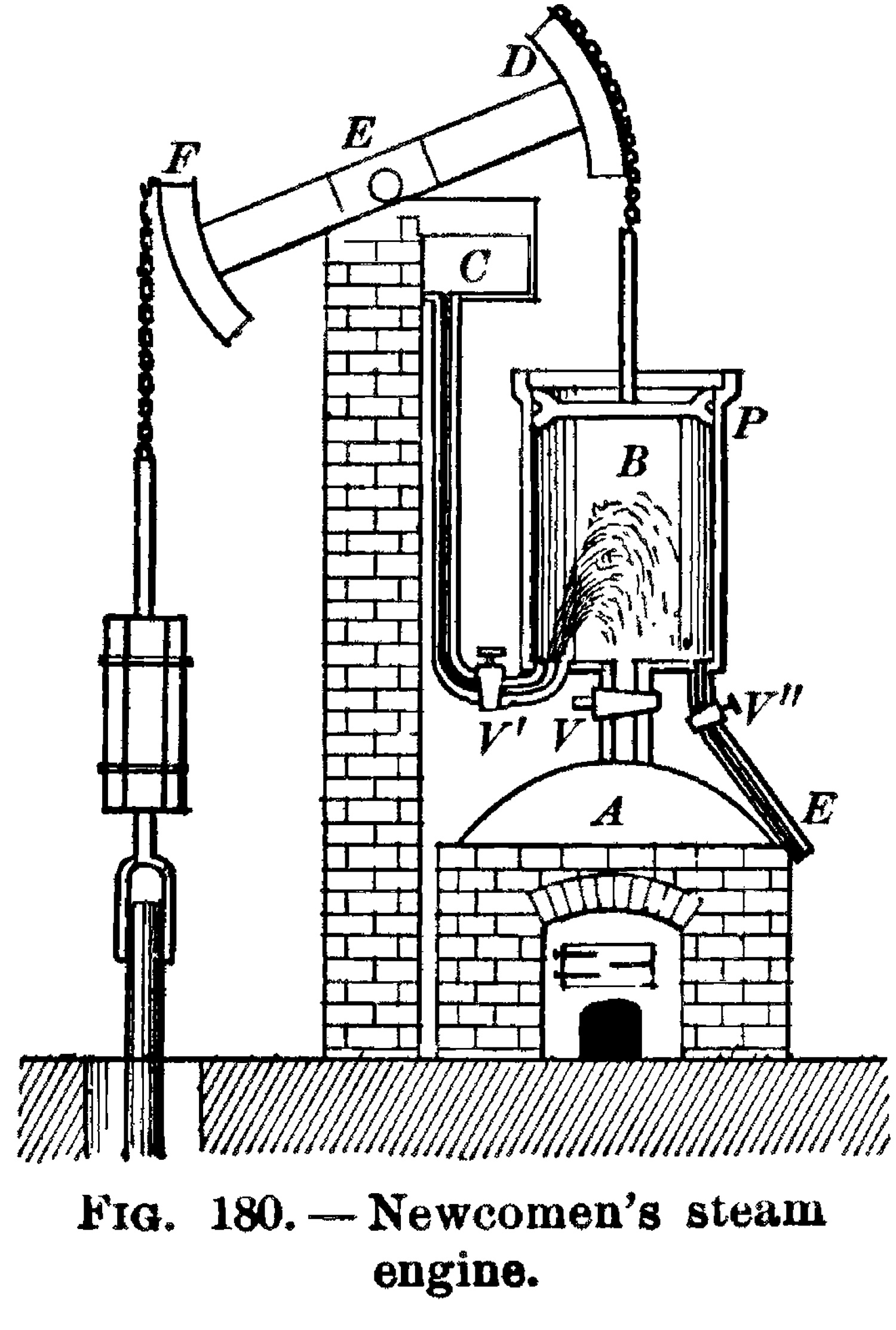 Паровой двигатель Ньюкомена, из книги Блэка и Дэвиса,
Практическая физика
, 1913
Паровой двигатель Ньюкомена, из книги Блэка и Дэвиса,
Практическая физика
, 1913
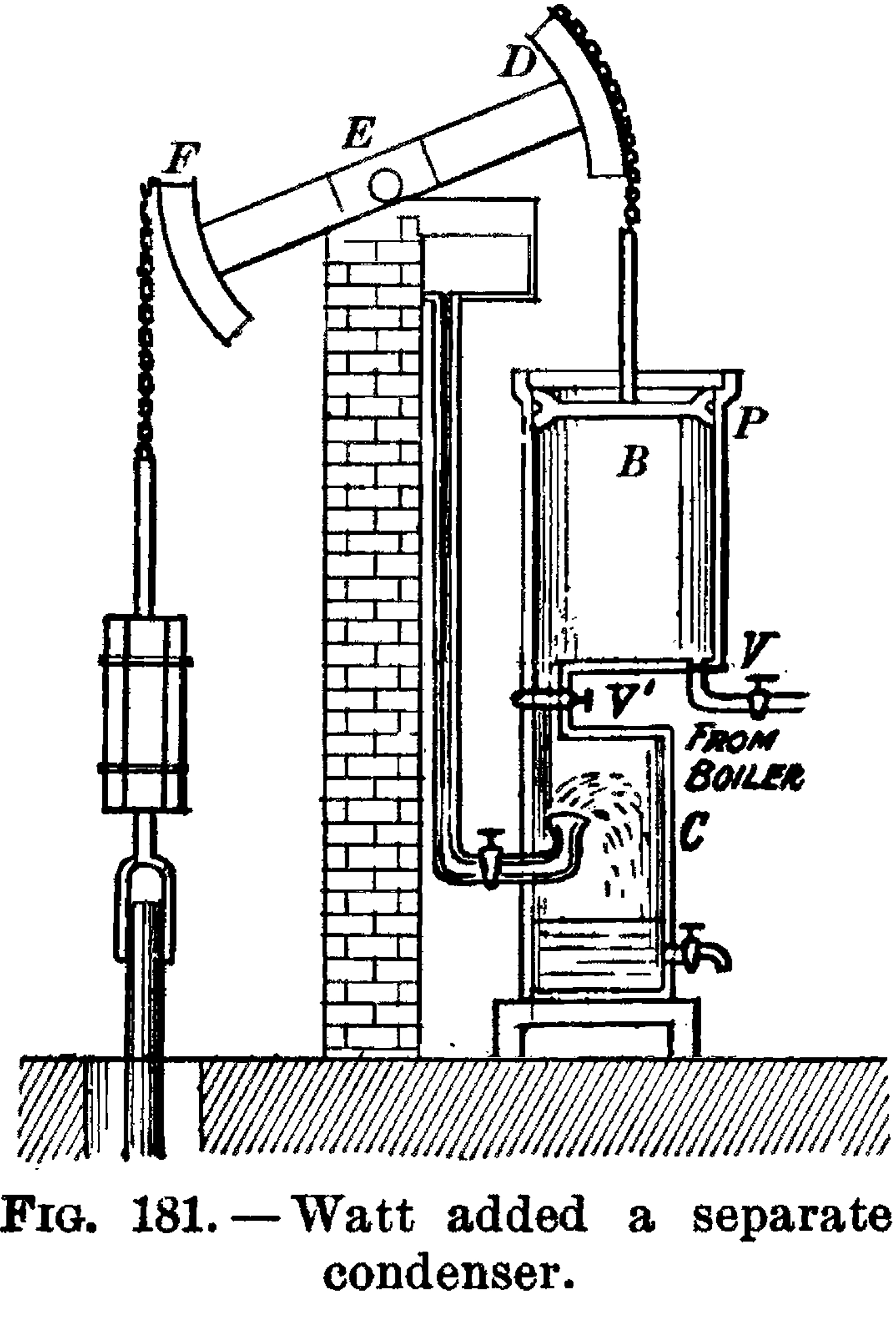 Паровой двигатель Уатта, там же.
Паровой двигатель Уатта, там же.
Скоро через этот искусственный метаболизм будет проходить гораздо больше энергии, чем через цикл Кребса в наших собственных телах. Это было, в широком смысле, о котором я говорю в этой книге, значительное эволюционное преобразование: симбиогенетическое событие между людьми и машинами, подобно более ранним симбиозам между людьми и тягловыми животными. Машинный метаболизм позволил человеческому обществу взорваться с прединдустриального уровня (около одного миллиарда человек в 1800 году, большинство из которых жили в крайней бедности) до современного масштаба (восемь миллиардов, большинство из которых больше не живут в бедности).
В процессе мы стали почти так же зависимы от машин для нашего существования, как они зависят от нас. Однако, даже когда угольные двигатели преобразовывали викторианский ландшафт — как в переносном, так и в буквальном смысле, ведь загрязнение было ужасным — никто не понимал их на глубоком уровне.
Что такое тепло и как его можно преобразовать в физическую работу? В течение некоторого времени ведущей теорией было представление о том, что тепло — это некое невидимое, невесомое вещество, «калорик», которое может таинственным образом проникать в другие материалы и проходить через них.
Объединив ньютоновскую физику, статистические расчеты и экспериментальные испытания, Больцман и его коллеги разобрались в том, что на самом деле происходит. Их концептуальная модель представляла собой трехмерную версию бильярдного стола, где шарики — это молекулы газа, мчащиеся по запечатанному пространству и отскакивающие от его стенок. Вычисление среднего эффекта всех этих отскоков привело к Закону идеального газа, который установил теоретические связи между давлением, объемом и температурой газа. Теория хорошо совпадала с наблюдениями экспериментаторов и инженеров.
Объем — это просто размер камеры, но идея о том, что давление — это совокупная сила на стенках камеры, когда молекулы отскакивают от них, а температура — это средняя кинетическая энергия этих мчащихся молекул, стала глубоким открытием. Не было необходимости в каком-либо таинственном «калорическом» веществе; тепло — это просто движение на микроскопическом уровне. А стремление к случайному распределению молекулярных положений и импульсов объясняет, почему, если открыть клапан между двумя камерами с газом при разных давлениях и/или температурах, эти давления и температуры быстро уравновесятся.
Прежде чем перейти к более сложным аспектам термодинамики, давайте добавим немного реализма в нашу вселенную бильярдных шаров. Мы знаем, что шарики, отскакивающие от бильярдного стола, не могут бесконечно продолжать свое движение. Они сталкиваются с трением, замедляясь при катании. И когда они сталкиваются друг с другом, столкновения немного «неупругие», что означает, что после столкновения они движутся немного медленнее, чем до него. Вскоре они останавливаются.
Как это возможно? На микроскопическом уровне законы физики обратимы. Импульс, как предполагается, должен сохраняться.
И количество материи и энергии также остается постоянным, независимо от того, движется ли время вперед или назад. Это Первый Закон термодинамики!
Приближаясь к реальному бильярдному столу, мы увидим, что шары — не самые мелкие элементы, сталкивающиеся друг с другом. Каждый бильярдный шар состоит из множества колеблющихся молекул, связанных между собой. Именно столкновения этих отдельных молекул заставляют шары отскакивать друг от друга — или, в данном случае, позволяют шару катиться по сукну, а не проваливаться сквозь него.
В каждом случае происходит передача импульса между молекулами. Каждый раз, когда это происходит, распределение молекулярных импульсов становится немного более случайным, то есть менее связанным с тем, в каком шаре находится молекула, или, в самом деле, находится ли она вообще в шаре или же в сукне под ним.
В самом случайном распределении молекулярных скоростей не будет никакой корреляции между скоростями двух молекул в одном шаре и скоростями молекул в разных шарах. Каждый шар будет незаметно дрожать на месте, при этом каждая из его молекул будет вносить крошечный вклад в этот танец. Мы называем это случайное дрожание «теплом». Когда все связанные движения превращаются в равномерно распределенное тепло, мы достигаем стабильного равновесия.
Пока шары все еще катятся, корреляции между скоростями их молекул отнюдь не равны; распределение далеко от случайного. Это не стабильное равновесие. Следовательно, неизбежность трения и неупругость столкновений — это статистические явления — просто еще одни симптомы неумолимого Второго Закона.
Итак, давайте снова представим, что бильярдные шары — это неделимые частицы, а не связанные коллекции молекул. В этом случае все столкновения должны быть упругими, все движения — без трения, а беспорядочно сталкивающиеся шары составят равновесие.
Как только система достигает равновесия, она остается в этом состоянии навсегда — конечное состояние, которое лорд Кельвин назвал «тепловой смертью». Это, казалось бы, тривиальное наблюдение имеет глубокие последствия. Во-первых, стрела времени теряет свое значение; любые два последовательных момента A и B могут быть упорядочены так же вероятно как B и A. Таким образом, невозможно сказать, что является причиной, а что следствием.
Связано с этим и то, что никакая работа не может быть выполнена. Если бы система находилась вне равновесия — например, если бы все шары находились с одной стороны бильярдного стола — мы могли бы установить подвижную преграду между пустой и занятой сторонами, соединенную с нагруженным коленчатым валом. Когда шары будут отскакивать, они будут толкать преграду, выполняя работу. То, что я только что описал, конечно, является поршнем, как в паровом двигателе.
Но если шары равновероятно могут находиться в любом месте, то, независимо от того, как быстро они движутся и отскакивают, не будет места для установки преграды, которое привело бы к какой-либо результирующей силе. Поршень не будет двигаться, потому что будет подвергаться одинаковому воздействию со всех сторон. Эта идея может быть обобщена: работа может быть выполнена только системой в неравновесии, например, когда давление или температура высоки в одном месте и низки в другом. Вот почему двигатель Ньюкомена нуждался и в горячем паре, и в холодной воде.
Я уже использовал термин «свободная энергия», но теперь мы можем его определить. Свободная энергия системы — это количество работы, которую она может выполнить. Далеко от равновесия, когда энтропия низка, большая часть кинетической энергии в бильярдных шарах является «свободной»; она может быть использована для движения поршней, подъема грузов, производства электрических токов, выполнения вычислений или запуска метаболических процессов. Но в состоянии равновесия энтропия максимальна, и свободная энергия равна нулю.
Это понимание взаимосвязи между энергией, энтропией и работой лежит в основе термодинамики — и жизни.
Динамическая стабильность
Помните, что жизнь казалась Шрёдингеру глубоко странной, потому что живые существа, по всей видимости, нарушают Второй закон термодинамики. Если бактерия, которую мы помещаем в колбу с водой, жива, а не мертва, если свободная энергия доступна в форме, которую бактерия может использовать, и вода содержит простые молекулы, подходящие для создания новых бактерий, то со временем мы увидим прямо противоположное увеличению беспорядка. Через некоторое время колба будет полна бактерий, которые размножаются, сотрудничают и конкурируют друг с другом.
Бактерии даже будут эволюционировать. Если колба достаточно велика — например, размером с планету — и мы подождем несколько миллиардов лет, то в ней могут оказаться существа столь же сложные, как мы, вместе с городами, развитыми технологиями и, возможно, планами по колонизации ближайшей колбы.
Ни один из этих процессов не может происходить без свободной энергии. Для нас она в первую очередь поступает от солнца. Теродинамика говорит нам, что даже если Второй закон, кажется, нарушается локально, он все равно действует, когда мы смотрим на ситуацию в более широком масштабе. Порядок, созданный в одном месте, обходится увеличением беспорядка в другом. Отсюда и загрязнение, и конечный срок жизни солнца, и неизбежная тепловая смерть вселенной.
Что нас здесь беспокоит, так это не эта общая картина, а ее кажущиеся локальные нарушения и то, как они, похоже, становятся все более трансгрессивными с течением времени. Загадка заключается не только в том, что бактерии существуют, но и в том, что с течением времени жизнь на Земле, кажется, становится все более сложной: от прокариотов к эукариотам; от эукариотов к многоклеточным животным; от простых многоклеточных животных к тем, у кого есть нервные системы; от умных животных к сложным обществам; от лошадей и плугов до космических путешествий и искусственного интеллекта.
Существует ли какой-либо общий принцип, стоящий за процессом усложнения, нечто вроде «однако» или «да, и» к мрачному Второму закону? И может ли он объяснить не только эволюцию и усложнение, но и абиогенез?
Да, и да. Модель bff может предложить нам сильно упрощенную систему для понимания этого принципа, так же как идеализированный бильярдный стол помогает нам осознать основы термодинамики.
Репликаторы возникают в bff, потому что сущность, которая воспроизводится, более «динамически устойчива», чем та, которая этого не делает. Другими словами, если мы начнем с одной ленты, которая может воспроизводиться, и одной, которая не может, то через некоторое время мы, скорее всего, найдем много копий той, что может воспроизводиться, но вряд ли наткнемся на другую, так как она будет разрушена шумом или перезаписана.
Эдди Просс, профессор-эмерит химии в Университете Бен-Гуриона в Негеве, описывает тот же феномен, используя более громоздкое выражение «динамическая кинетическая стабильность» (ДКС).
Я опущу «кинетическую», поскольку эта идея также применима далеко за пределами области «химической кинетики» Просса (описывающей скорости химических реакций). В bff, например, динамическая стабильность может также относиться к программам или фрагментам программ.
Как указывает Просс, популяция реплицирующихся молекул может быть более стабильной, чем даже самые стойкие из пассивных материалов. Пассивный объект может быть хрупким, как мыльный пузырь, или прочным, как каменная скульптура. Скульптура может прослужить дольше, но в конечном итоге она все равно эфемерна. Каждое взаимодействие с чем-либо в мире приведет к деградации ее состава или структуры, ее индивидуальная идентичность начнет размываться. Для скульптуры все идет под откос. Это и есть работа Второго закона, как обычно.
Самовоспроизводящаяся молекула — как ДНК внутри живой бактерии — это совсем другое дело. Она термодинамически хрупка, особенно если мы считаем, что ее идентичность состоит не только в общей структуре, но и в длинной последовательности специфических нуклеотидов.
Однако его структура не просто устойчива, но и «антихрупка». Пока ДНК способна к воспроизводству — это по своей сути динамический процесс — эта структура может существовать практически вечно. Небольшой стресс или неблагоприятные условия могут даже помочь ДНК сохранить или улучшить свою функциональность. Вот как порядок преодолевает хаос. На самом деле, дарвиновский отбор эквивалентен Второму Закону, если мы расширим наше представление о стабильности, чтобы включить популяции репликаторов. С термодинамической точки зрения, центральное наблюдение Дарвина заключалось в том, что более эффективный репликатор стабильнее, чем менее эффективный. Как говорит Просс, [М]атерия […] имеет тенденцию трансформироваться […] из менее стабильных форм в более стабильные. […] [Э]то и есть суть химической кинетики и термодинамики […]. А какой центральный закон управляет такими преобразованиями? Второй Закон. […] В обоих [статическом и кинетическом] мирах химические системы стремятся трансформироваться в более стабильные […] — термодинамическая стабильность в «обычном» химическом мире, динамическая кинетическая стабильность в мире репликаторов. Как химик, Просс чувствителен к тесным связям между энергией, энтропией и стабильностью, будь то статической или динамической. Однако он не устанавливает явной связи с теорией вычислений. Теперь кажется очевидным, что, объединив термодинамику с теорией вычислений, мы сможем понять жизнь как предсказуемый результат статистического процесса, а не воспринимать её с тревогой как технически допустимую, но таинственную. Наши эксперименты с искусственной жизнью показывают, что, когда вычисление возможно, оно будет «динамическим аттрактором», поскольку реплицирующие сущности более динамически стабильны, чем нереплицирующие; и, как показал фон Нейман, репликаторы по своей сути являются вычислительными. Bff не имеет понятия о энергии, но в нашей вселенной репликаторы требуют источник энергии.
Это происходит потому, что вычисления в целом включают в себя необратимые шаги — иначе говоря, причины и следствия — и, следовательно, вычисления потребляют свободную энергию. Вот почему чипы в наших компьютерах потребляют электроэнергию и выделяют тепло во время работы. (И почему мой компьютер нагревается, когда я запускаю bff.) Жизнь также должна потреблять энергию и выделять тепло по той же причине: она по своей сути вычислительна.
Комплексизация
Когда мы извлекаем ленту из супа bff после миллионов взаимодействий, когда репликаторы уже захватили инициативу, мы часто наблюдаем уровень сложности в программе на этой ленте, который кажется ненужным — даже неправдоподобно высоким. Рабочий репликатор мог бы состоять всего из нескольких инструкций в одном цикле, требуя всего лишь пару сотен операций для выполнения. Вместо этого мы часто видим инструкции, заполняющие большую часть из 64 байт, множество сложных вложенных циклов и тысячи операций на каждое взаимодействие.
Откуда взялась вся эта сложность? Она определенно не выглядит как результат простого дарвиновского отбора, действующего на случайный текст, сгенерированный пословно миллионом обезьян, печатающих на миллионе пишущих машинок.
На самом деле такая сложность возникает даже при нулевой случайной мутации — то есть, учитывая только начальную случайность в супе, которая составляет меньше байт, чем текст этой книги. Вряд ли это миллион обезьян — и слишком мало случайных байтов, чтобы содержать более чем несколько последовательных инструкций, не говоря уже о целой рабочей программе.
Ответ напоминает о великом прозорливом выводе Линн Маргулис: центральной роли симбиоза в эволюции, а не случайной мутации и отбора. Когда мы внимательно смотрим на период покоя в супе bff перед началом репликации лент, мы замечаем устойчивый рост объема вычислений. Мы наблюдаем быстрое возникновение несовершенных репликаторов — очень коротких фрагментов кода, которые так или иначе имеют некоторую ненулевую вероятность генерации большего количества кода.
Даже если созданный код отличается от оригинала, он все равно остается кодом, и только код может порождать новый код; некод не способен на это! Таким образом, с самого начала действует процесс отбора, в котором код порождает код. Этот по своей сути творческий, самокатализирующийся процесс гораздо важнее случайных мутаций в создании новизны. Когда фрагменты размножающегося кода объединяются, чтобы сформировать репликатор, это симбиотическое событие: работая вместе, эти кусочки кода генерируют больше кода, чем могли бы по отдельности, и созданный ими код, в свою очередь, произведет еще больше кода, который сделает то же самое, в конечном итоге приводя к репликации целой ленты и экспоненциальному росту.
Ближе к супу bff перед экспоненциальным ростом можно выделить различные фазы "преджизни", которые могли иметь близкие аналоги в процессе абиогенеза на Земле. На первой фазе отдельные инструкции время от времени генерируют другую индивидуальную инструкцию, но это скорее похоже на простую химическую реакцию, чем на настоящие вычисления; инструкции не действуют как часть какой-либо более крупной программы.
На второй фазе мы начинаем видеть инструкции в определенных позициях или в определенных комбинациях, которые с большей вероятностью ведут к копиям самих себя, чем можно было бы ожидать случайным образом, хотя часто и косвенно. Начинают формироваться "автокаталитические наборы": циклы динамических взаимодействий, которые взаимно усиливают друг друга. Эти наборы также могут возникать спонтанно в химическом мире и давно предполагается, что они способствовали абиогенезу.
На этом этапе, с размножающимися и сталкивающимися автокаталитическими фрагментами кода, становится возможен скачок, который выводит нас за пределы мира цифровой химии и в мир настоящих вычислений: появление первых истинных репликаторов. Это уже не просто автокаталитические наборы, а короткие программы, которые копируют себя или друг друга, используя циклические инструкции.
С этим переходом к вычислениям приходит огромное улучшение эволюционной способности, поскольку теперь любое изменение, внесенное в программу, которое не нарушает ее цикл копирования, будет наследуемым. Таким образом, классический дарвиновский отбор может вступить в действие, позволяя адаптироваться к меняющейся среде или образовывать новые виды для занятия различных ниш. Если мы настаиваем на различии между неживой и живой природой, это может быть разумным местом для проведения этой границы.
Тем не менее, эти самые ранние реплицирующие программы вряд ли смогут аккуратно скопировать целую ленту. Обычно они копируют лишь короткие участки ленты, которые могут вставляться в произвольные места, что приводит к непредсказуемым результатам. По мере того как эти лоскутные, фрагментарные репликаторы размножаются, бфф-суп переходит в хаотическую фазу. Несмотря на эту неразбериху, на данном этапе ленты еще не приобрели очевидной структуры. На первый взгляд они по-прежнему выглядят как случайный мусор, хотя растущее количество вычислений (измеряемое как среднее количество операций на взаимодействие и плотность инструкций на ленте) указывает на то, что что-то происходит.
Отслеживание происхождения каждого байта в супе, начиная с случайной инициализации, может помочь разобраться в кажущемся хаосе. Сначала почти каждый байт каждой ленты остается тем, чем он был в момент инициализации; если провести линию от текущего положения каждого байта на ленте к его исходной позиции, линии будут тянуться назад к нулевому времени параллельно, как нити утка на ткацком станке. Время от времени байт изменяется, перерезая нить, или копируется, перемещая ее по диагонали относительно других нитей.
▶ Происхождение отдельных байтов на лентах в бфф-супе после 10 000, 500 000, 1.5 миллиона, 2.5 миллиона, 3.5 миллиона, 6 миллионов, 7 миллионов и 10 миллионов взаимодействий. Растущая роль самосмещения в создании новизны очевидна, достигая своего пика (незадолго до 6 миллионов взаимодействий) с появлением репликатора, способного копировать целую ленту, части которого являются модифицированными копиями более короткого несовершенного репликатора.
С появлением неукротимых программ-репликаторов все оставшиеся зависимости от прошлого быстро исчезают, когда репликаторы в неистовом порыве творческого разрушения копируют друг друга. Каждый отдельный байт может быть скопирован сотни раз на различные участки ленты, в процессе стирая все, что было там ранее. Вскоре все эти копии могут быть уничтожены другим, более эффективно реплицирующим фрагментом. В результате история каждого байта становится очень краткой по времени, но сложной по пространству — короткий, но запутанный клубок боковых скачков. Ткачество становится чистым утком, без основы.
Соревнуются ли эти неукротимые репликаторы или сотрудничают? И то, и другое. Репликаторы, которые не могут справиться с нагрузкой, уничтожаются вместе с любыми нереплицирующими байтами. В то же время выжившие репликаторы постоянно образуют химеры, рекомбинируясь с другими репликаторами (или даже копиями самих себя), чтобы стать еще более эффективными. Это снова симбиогенетические события: под-единицы сливаются, образуя более крупную и способную супер-единицу.
Эта хаотичная фаза является таким мощным тиглем для направленной эволюции, что, как правило, не длится долго. Она быстро производит надежный репликатор целой ленты, который затем начинает экспоненциально развиваться, что приводит к драматическому переходу к организованным структурам (и большому количеству вычислений), которые являются наиболее очевидной чертой bff. В этот момент искусственная жизнь кажется спонтанно возникающей.
Но, как мы теперь можем понять, в этом нет ничего спонтанного. Репликаторы были здесь все это время, и каждый более крупный репликатор состоит из меньших — перевернутое дерево жизни, состоящее из слияний с течением времени, а не из делений.
Тем не менее, творческая работа эволюции еще не завершена. После взлета полностью функционального репликатора целой ленты мы часто наблюдаем дальнейшие симбиотические события. С классической дарвиновской точки зрения это кажется загадочным, поскольку дальнейшая эволюция кажется излишней, когда целые ленты надежно реплицируются.
Как же "фитнес" может улучшаться дальше, если каждая лента полностью копирует себя при каждом взаимодействии с другой лентой?
Необходимо учитывать, что поскольку инструкции для репликации целой ленты не занимают все 64 байта, на ленте остается свободное место, которое можно посвятить... чему угодно. В этом и заключается суть репликации в стиле фон Неймана — она позволяет открытой эволюции, поскольку лента может содержать дополнительную информацию, помимо кода, необходимого для самой репликации.
Любые дополнительные реплицированные байты могут, конечно, быть случайными — просто пассивный, бессмысленный информационный груз, который путешествует от одного поколения к другому. Но если эти байты содержат инструкции, то эти инструкции могут выполняться. А если они могут выполняться, то они тоже могут реплицировать себя. Таким образом, симбиогенетический процесс может продолжать функционировать, создавая дополнительные репликаторы внутри уже реплицирующейся ленты. Иногда эти субрепликаторы даже производят несколько копий самих себя за одно взаимодействие.
Субрепликаторы могут взаимодействовать со своим хозяином различными способами. Они могут "убить" хозяина, повредив его код репликации, что обычно катастрофично для субрепликатора, так как тем самым он разрушает среду, в которой может функционировать. Субрепликаторы могут быть нейтральными, оставляя механизмы репликации хозяина в покое. Или они могут быть симбиотическими, например, обеспечивая устойчивость к мутационным повреждениям за счет избыточного копирования кода хозяина. Общая тенденция направлена к симбиозу, так как это наиболее динамично стабильное состояние.
Со временем код колонизирует значительную часть из 64 байтов. Код более динамично стабилен, чем не код, и его динамическая стабильность увеличивается через симбиоз с еще большим количеством кода — в частности, когда фрагменты кода находят способы работать в функциональном тандеме.
гистограмма, показывающая, сколько из 64 байтов ленты кодируют инструкции; обратите внимание, как и прежде, на продолжающийся рост после полной репликации ленты, который здесь начинается примерно с 3 миллионов взаимодействий.
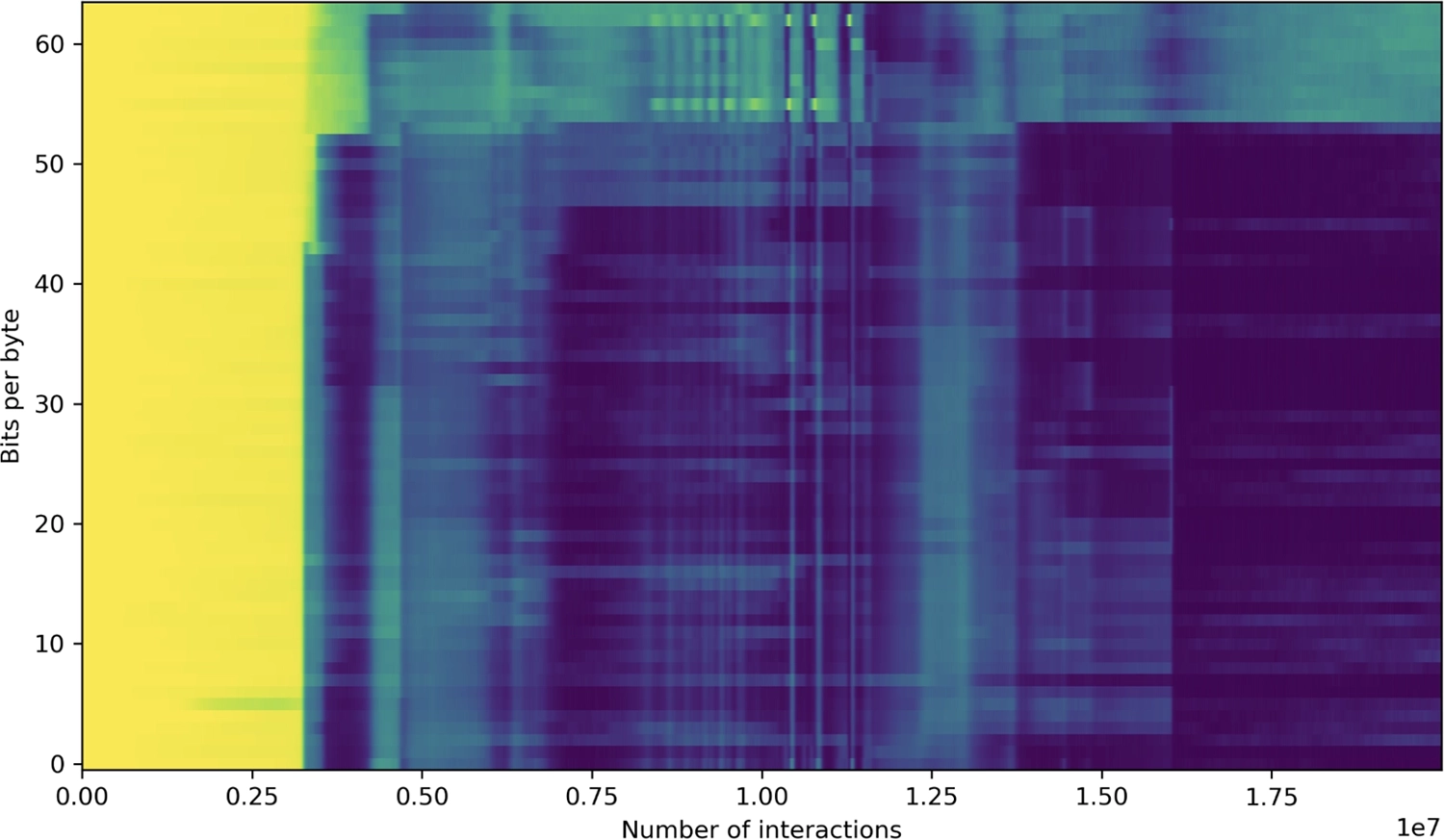
Переменность или количество информации в байте ленты на каждой позиции, выраженное в энтропии в битах. Перед переходом значение близко к 8 (что соответствует случайному распределению значений от 0 до 255), но в момент перехода к полной репликации ленты оно резко падает; однако некоторые позиции более переменны, чем другие, с большей переменностью к концу ленты.
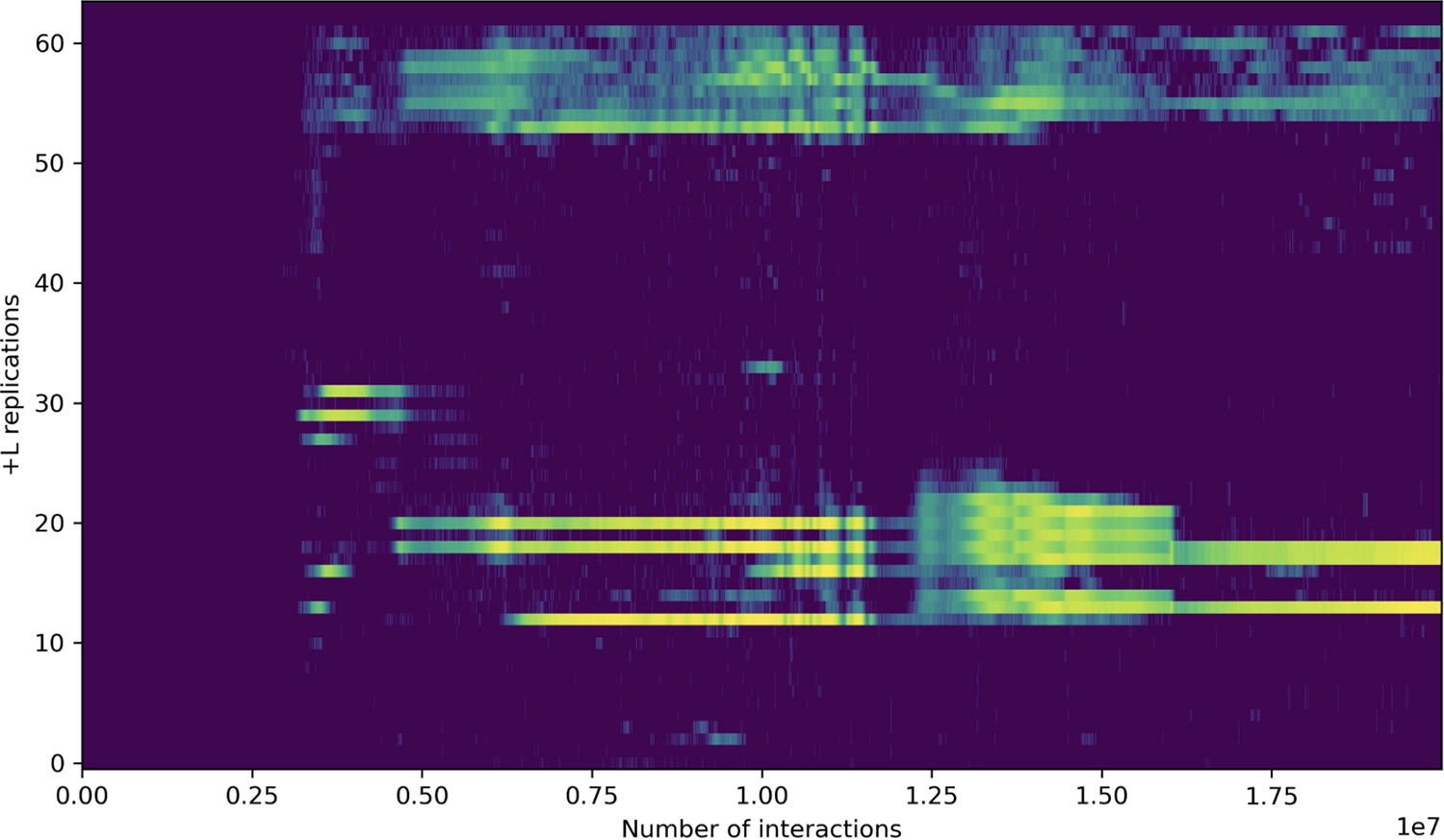
Гистограмма, показывающая позиции инструкций цикла “[”, которые выполняют «выравненную» копию области ленты с одной ленты на ту же позицию на другой ленте, что соответствует идеальной репликации. Обратите внимание, что в «супе» есть несколько идеальных репликаторов, популяции которых меняются со временем.
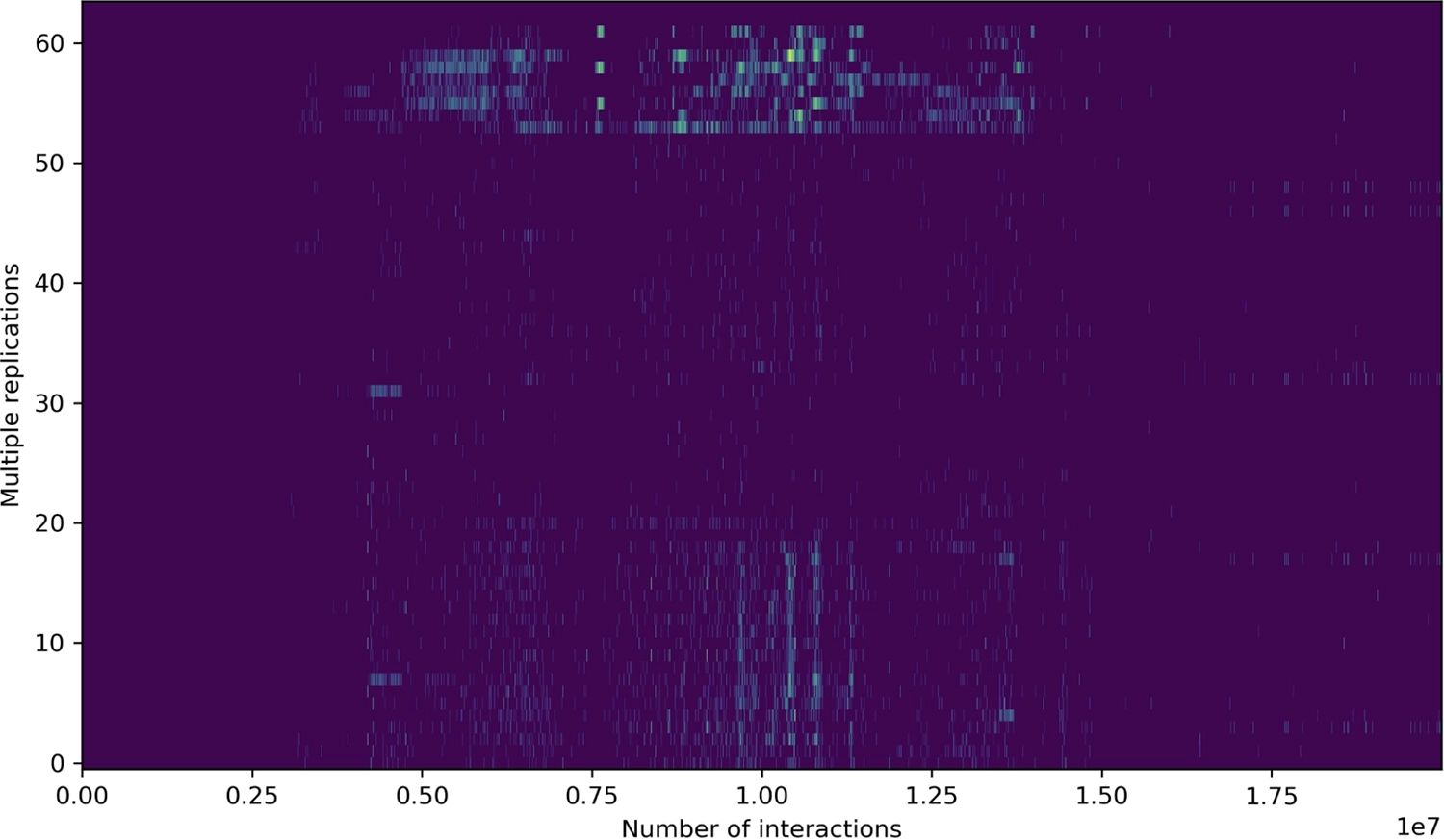
Гистограмма, показывающая позиции инструкций цикла “[”, которые реплицируют себя несколько раз при выполнении, появляясь в этом запуске после полной репликации ленты.
В некотором смысле симбиоз — это сама суть функциональности. Когда мы говорим о функции почки, которая имеет смысл только в контексте, мы имеем в виду, что она находится в симбиозе с другими функциями — такими как функции печени (разложение аммиака на мочевину), сердца (качание крови) и так далее. Каждая из этих функций целенаправленна именно потому, что ее входные данные являются выходными данными других, а ее выходные данные — входными данными для других, и таким образом они формируют сеть динамически стабильных циклов.
То же самое верно и для более крупных, планетарных взаимосвязей. «Цель» растений, с точки зрения животной жизни, заключается в производстве кислорода и сахара, которые мы дышим и едим. «Цель» животных, с точки зрения растений, состоит в том, чтобы превращать кислород обратно в углекислый газ, а также обеспечивать компост и опыление.
Наше растущее понимание жизни как самоподдерживающегося динамического процесса сводится не к вещам, а к сетям взаимовыгодных отношений. На каждом уровне жизнь представляет собой экологию функций. Поскольку функции могут быть выражены в вычислительном виде, мы могли бы даже сказать, что жизнь — это код. Индивидуальные вычислительные инструкции являются неразрывными кванты жизни — минимальный набор сущностей, которые, как бы нематериальными и абстрактными они ни казались, объединяются, чтобы сформировать более крупные, более стабильные и более сложные репликаторы в постоянно восходящих симбиотических каскадах.
В игрушечной вселенной bff семь специальных символов “<>+−,[]” являются элементарными инструкциями. На примитивном дне океана геотермально управляемые химические реакции, способные катализировать дальнейшие химические реакции, вероятно, играли ту же роль. В других условиях, на другой планете или в другой вселенной, многие различные элементарные взаимодействия могли бы выполнять ту же функцию — лишь бы они были Тьюринг-полными, что позволяло бы им сделать скачок от автокаталитических наборов к истинной репликации.
Вирусность
Хотя bff — это всего лишь игрушечная вселенная, она может служить упрощенной моделью жизни и эволюции, так же как ньютоновская вселенная бильярдных шаров может служить упрощенной моделью термодинамики идеальных газов. Как мы уже видели, некоторые предсказания bff довольно сильно отличаются от классической дарвиновской теории:

Морское чудовище из "Истории чудовищ" Улисса Альдрованди, 1642
Bff предполагает, что симбиогенез является более важным двигателем эволюционных инноваций, чем случайная мутация. Поскольку симбиогенез должен включать комбинации уже существующих динамически стабильных сущностей, мы должны ожидать, что сложные реплицирующие сущности появятся после (и будут состоять из) более простых. В результате, углубляясь в субрепликаторы внутри более крупного репликатора, мы должны иметь возможность заглянуть назад в эволюционное время.
Эти явления должны показаться вам знакомыми! Они полностью соответствуют наблюдениям Линн Маргулис, которые противоречили биологической ортодоксии двадцатого века, но теперь, если не стали мейнстримом, то, по крайней мере, начинают завоевывать уважение.
Но это еще не все:
Из-за нестабильности несовершенных репликаторов, предшествовавших первому истинному репликатору, мы можем ожидать, что этот первый истинный репликатор станет историческим «горизонтом событий», задавая шаблон для всего последующего и стирая независимые следы того, что было до него.
Мы наблюдаем это и на Земле. Хотя первые химические шаги к протожизни могут все еще происходить в таких средах, как черные курильщики, мы не видим недостающих звеньев. Где же те несовершенные репликаторы, которые объединились, чтобы образовать самые простые формы жизни сегодня?
Скорее всего, мы не видим их в одиночку, потому что, как и в случае с bff, их нестабильность сделала их эфемерными, быстро вытесненными или поглощенными первой стабильно реплицирующейся клеткой — которая, возможно, уже была узнаваемо родственной современным бактериям и археям. (Эволюционные биологи называют это «Последним универсальным общим предком» или LUCA.) Тем не менее, мы можем предположить, что многие из составных частей самых древних сохранившихся форм жизни — таких как обратный цикл Кребса и репликация РНК — являются окаменелыми фрагментами более ранних несовершенных репликаторов.
В том же духе:
Эволюционный код должен не только содержать инструкции для репликации самого себя в целом, но и быть насыщенным подпоследовательностями, которые содержат инструкции для независимой репликации самих себя.
Если симбиоз между этими частями порождал новизну, движущую эволюцией целого, мы должны увидеть доказательства в геноме многих «сломанных» или неполных субрепликаторов.
Код, который эволюционировал через такую иерархическую симбиотическую репликацию, также должен содержать множество последовательностей, которые являются повторяющимися или копиями других частей.
Мы можем найти все эти особенности в нашем собственном генетическом коде — и они не соответствуют тому, что мы ожидали бы увидеть, если бы наши геномы эволюционировали в основном через мутации и отбор. Когда в 2000 году была опубликована первая полная последовательность человеческого генома, нас ждали некоторые сюрпризы. Один из них — удивительно высокая доля так называемого «мусорного ДНК», которое не кодирует белки: около девяноста восьми процентов. Как оно туда попало? Действительно ли оно что-то делает? Теперь мы понимаем, что часть этого «мусора» участвует в регуляции генов, но большая часть все еще остается с неизвестной или неопределенной функцией. Однако при близком рассмотрении оказывается, что большая часть нашего кода, будь то «мусор» или нет, является… вирусной.
Относительные пропорции основных семейств человеческих геномных элементов, по Lander et al. 2001
Репродуктивный цикл вирусов — это часто рассказываемая история. Путешествуя между клетками, вирус выглядит как кусочек ДНК или РНК, упакованный в белковую «оболочку», способную связываться с целевой клеткой и вводить свой генетический груз. Оказавшись внутри клетки, вирусный код захватывает клеточные ресурсы, чтобы начать репликацию, копируя свой генетический материал и производя белки своей оболочки. Эти белки собираются в новые вирусы. Безумно производя вирусные белки, клетка в конечном итоге может разорваться, выпуская поток вирусных частиц, чтобы повторить цикл.
«Ретровирусы», такие как ВИЧ (вирус иммунодефицита человека, вызывающий СПИД), включают дополнительное «обратное транскрипционное» оборудование для постоянного встраивания своего генетического материала в ДНК клетки. Это делает их гораздо труднее для организма хозяина. Ключевой фермент, «обратная транскриптаза», был охарактеризован только в 1970 году, но Барбара МакКлинток (1902–1992), работая в лаборатории Cold Spring Harbor на Лонг-Айленде, сделала более раннее открытие, которое оказалось тесно связанным с этим.

Барбара МакКлинток, Кафедра генетики, Карнеги Институт в Колд-Спринг-Харбор, Нью-Йорк, 1947 год
Изучая генетику кукурузы в конце 1940-х и начале 1950-х годов, МакКлинток обнаружила подвижные сегменты ДНК, которые, казалось, могли «вырезать» и «вставлять» себя в разные участки генома, создавая разнообразные мозаичные цветовые узоры на зернах кукурузы.
Хотя многие из её коллег изначально скептически относились к этим «транспозонам» или «прыгающим генам», в конечном итоге МакКлинток была удостоена Нобелевской премии за свои исследования.
С тех пор было открыто гораздо большее разнообразие «транспозируемых элементов», некоторые из которых не просто «вырезают и вставляют», но и «копируют и вставляют» себя; они фактически являются репликаторами внутри нашей ДНК. На самом деле, некоторые из них представляют собой полноценные ретровирусы, содержащие инструкции не только для встраивания своего кода в геном клетки-хозяина, но и для производства оболочечных белков, позволяя этому коду выходить в мир в поисках других клеток для заражения.
Ретровирусы удивительно …
интимны.
ВИЧ специально нацеливается на иммунные клетки, но если ретровирус заражает яйцеклетку или сперматозоид, он может вставить свой код в зародышевую линию организма, становясь постоянной частью не только этой клетки, но и целого вида. В публикации 2006 года в престижном журнале
Nature
исследователи из Университета Квинсленда в Австралии сообщили о том, что впервые поймали ретровирус в момент его превращения в «эндогенный».
Эпидемия лейкемии у коал изначально была связана с ретровирусом, а затем было установлено, что этот ретровирус вторгся в зародышевую линию коал за предыдущий век, что сделало болезнь наследственной.
Означает ли это конец для коал? Вероятно, нет.
Человеческий геном полон признаков того, что тот же процесс происходил много раз в истории нашего вида.
По меньшей мере восемь процентов нашего генома составляют эндогенизированные ретровирусы — остатки таких вирусных инвазий. Помните, восемь процентов — это в несколько раз больше ДНК, чем кодирует наши «собственные» гены!
Мы обычно воспринимаем вирусы как простые агенты болезней, которые паразитируют на наших клетках, чтобы размножаться, так как у них нет собственного репродуктивного механизма. Однако реальность может быть совершенно иной. Как многие из нас знают из недавнего опыта с COVID, вирусы (и патогены в целом) со временем склонны эволюционировать в менее смертоносные формы для своих хозяев — по очевидным причинам: мертвый хозяин — это тупик и для патогена. Существуют все более убедительные доказательства того, что отношения между вирусом и хозяином могут заходить еще дальше, становясь симбиотическими. Например, ретровирусный код в нашем геноме стал основополагающим для формирования плаценты, иммунной системы, дифференцировки клеток и функционирования мозга.
Более того, эндогенные ретровирусы — это лишь верхушка айсберга. Почти половина нашего генома состоит из транспозируемых элементов различного рода. Некоторые из них, вероятно, когда-то были ретровирусами, или наоборот. Полностью секвенированные геномы часто содержат длинные участки высокорепетитивных последовательностей — от длинных и сложных до простых чередований двух или трех символов — свидетельство о том, что субрепликаторы вышли из-под контроля.
Наши геномы, другими словами, воспроизводятся не только целиком, но и включают работающие репродуктивные подсеквенции на многих уровнях. Некоторые из них новые и могут все еще вызывать болезни; другие старше и, возможно, утратили способность образовывать вирусные оболочки или другие механизмы, необходимые для инфекционности; а некоторые интегрировались в наш собственный код так глубоко, что больше не являются отдельными. Часть этого кода даже выполняет критически важные функции в нашем организме. Очевидно, что, по крайней мере на последнем этапе этого процесса, установилась полная симбиоз: ни хозяин, ни субрепликатор не смогли бы выжить, не говоря уже о размножении, самостоятельно.
В своей книге 2009 года «Виролюция» автор и врач Фрэнк Райан утверждает, что вирусы, возможно, всегда были симбионтами. Вирусы чумы, такие как ВИЧ, не возникают из ниоткуда; они способны перескакивать между видами. Мы знаем, что ВИЧ изначально был ВИПЧ — вирусом иммунодефицита обезьян, разновидности которого встречаются у приматов Старого Света, включая африканских зеленых обезьян, черных мангобеев, мандрилов и шимпанзе. Однако многие из этих вирусов не причиняют вреда своим первоначальным хозяевам.
Также известно, что если вирус попадает в организм, физиология которого слишком отличается от физиологии его оригинального хозяина, он не может закрепиться — что, безусловно, является удачей для вас, если вы когда-либо глотали глоток морской воды, в которой, вероятно, содержалось около миллиарда вирусных частиц! (Большинство из них нацеливались бы на одноклеточные морские организмы.) Наибольшую опасность представляют вирусы, адаптированные к другому, но близкородственному виду. Возможно, когда такой вирус убивает, он выполняет свою задачу: уничтожает соперников, которые вторглись на территорию оригинального хозяина.
Вирусы, другими словами, могут работать как иммунная система вне тела. Внутри наших тел наши иммунные системы ищут и уничтожают клетки, которые распознаются как «не наши». Снаружи наши вирусы могут аналогичным образом искать и уничтожать целых животных, которые распознаются как «не наши». Однако, как только вирус становится эндемичным для новой популяции, даже если изначально он убивает многих, он будет дифференцироваться, соадаптироваться и, возможно, в конечном итоге станет местным.
Можно рассмотреть бактериальные патогены через аналогичную призму; так, хорошо задокументированные эпидемии оспы, которые европейские колонисты принесли в Америку, опустошили коренные популяции, а также вирулентная (хотя менее смертоносная) эпидемия сифилиса, которую, как считается, привез в Европу Колумб.
В отличие от бактерий, однако, ретровирусы не просто оседают в нашей среде или в наших телах, но сливаются с нашими геномами, становясь неотъемлемой частью нас — особенно когда они изменяют зародышевую линию.
По своему объему наша ДНК, похоже, состоит в основном из многослойных остатков множества таких прошлых слияний. Ясно одно: транспозируемые элементы и «эндогенные вирусные элементы» (EVEs) внесли гораздо больше изменений в наш геном за последние этапы эволюции, чем мутации.
И они занимаются этим уже очень долго. Судя по имеющимся данным, вирусы существуют как минимум с эпохи Последнего Универсального Общего Предка, если не дольше.
Эта непрекращающаяся экология подвижных, самовоспроизводящихся и даже инфекционных генов заставляет предполагаемое «дерево жизни» продолжать запутываться в самых странных формах. Например, целая четверть генома коровы состоит из копий ретротранспозона BovB. Похоже, что BovB многократно перескакивал между видами, создавая собственное филогенетическое дерево — «странную параллельную вселенную, где коровы ближе к змеям, чем к слонам, и где один геккон ближе к лошадям, чем к другим ящерицам».
Это примеры «горизонтального переноса генов» (HGT), долго ассоциировавшегося с бактериями, но явно более универсального.
Карта распространения ретротранспозонов BovB и L1 среди 759 видов эукариот, по данным Ivancevic et al. 2018
То, действует ли новый фрагмент ДНК в данный момент как друг, враг или что-то среднее, зависит от того, на каком этапе симбиотической траектории он находится. Когда он только появляется, вряд ли он будет дружелюбным. Но если он существует долго, и особенно если стал эндогенным, его динамическая стабильность подразумевает, что он станет неопасным или даже дружелюбным благодаря совместным эволюционным давлениям на хозяина, захватчика и «хозяина плюс захватчик» как «холобионт» или симбиотически слитое целое. Если какой-либо ген или другая функциональность в слиянии окажется полезной для большего целого, эта функциональность будет сохранена, даже если репродуктивные способности подрепликатора ухудшаются и в конечном итоге исчезают.
Возможно, именно так в целом и возникает новая генетическая функциональность. В этом свете случайная точечная мутация может рассматриваться как нечто вроде минимального события абиогенеза, создающего крошечную паразитическую «форму жизни» внутри генома. Самостоятельно она, конечно, не сможет воспроизводиться, так как не обладает независимыми репродуктивными механизмами; однако, как и вирус, мутация может воспроизводиться — используя ресурсы хозяина, который будет копировать её вместе со всем остальным. Когда такая крошечная, случайная сущность попадает в геном сложного существующего организма, она вряд ли окажется дружелюбной. Как и при любом более крупном вторжении, мутация либо повредит или убьёт своего хозяина (и себя вместе с ним), либо достигнет динамической стабильности, будучи нейтральной или (иногда) полезной.
Таким образом, так же как термодинамика репликаторов включает в себя классическую термодинамику как частный случай, эволюция через симбиогенез может охватывать классическую дарвиновскую теорию как частный случай. Точечная мутация вряд ли станет основным двигателем эволюции, когда жизнь уже начала развиваться, поскольку она значительно слабее и медленнее по сравнению с симбиогенезом более высокого порядка. В отличие от точечной мутации, фрагмент кода, который уже циркулировал, перемещаясь по геному или даже между видами, не является случайным. Он обязательно включает в себя реальную функциональность. И, по крайней мере в некоторых условиях, эта функциональность находилась под эволюционным давлением, чтобы помогать или, по крайней мере, не убивать своего хозяина. Эволюция набирает обороты со временем, и крупные эволюционные переходы становятся всё более частыми именно потому, что становятся возможными симбиогенезы всё более высокого порядка.
Сжатие
Когда код эволюционирует через симбиогенез, он приобретает любопытную статистическую структуру: его части будут копиями (или почти копиями) других частей, и по мере того как эти части устанавливают симбиоз, они образуют более крупный агрегат, который также будет копировать себя как единое целое.
Это напоминает (хотя и не совсем то же самое) «фрактал»: структуру, которая повторяет себя на каскаде все более мелких масштабов. Давайте сделаем небольшой крюк через фракталы и их связь с биологией, чтобы затем рассмотреть последствия того, что эволюция сама по себе проявляет определенные фрактальные свойства.
Франко-американский математик Бенуа Мандельброт (1924–2010) — наиболее известная фигура, ассоциирующаяся с фракталами, хотя они были впервые описаны английским метеорологом Льюисом Фраем Ричардсоном (1881–1953). После Второй мировой войны Ричардсон, квакер по вероисповеданию, пытался создать математическую модель для предсказания вероятности конфликтов между странами. Одним из параметров в его модели была длина их общей границы. Однако он был озадачен тем, что некоторые границы, похоже, не имели согласованной длины. Португальцы, например, утверждали, что их граница со Испанией составляет 613 миль, но испанцы настаивали, что она равна 754 милям. Это казалось странно большим расхождением для того, что должно было быть легко измеряемым расстоянием!
В конечном итоге Ричардсон разобрался в ситуации: испанцы использовали карту с более высоким разрешением. Каждое изгиб и поворот, которые португальцы могли видеть на реках Минью, Дору и Гвадиане, имели более мелкие изгибы и повороты, видимые только на испанской карте. Ричардсон показал, что если при увеличении любого извива открываются новые извивки, то общая длина «самоподобной» кривой, такой как береговая линия или река, может расти без ограничений, если измерять ее все меньшей линейкой.
Береговая линия Британии, измеренная с помощью линейок разной длины
Таким образом, «фрактальная геометрия» очень отличается от евклидовой геометрии, которую мы все изучали в школе, где длина линии или кривой четко определена и не увеличивается при увеличении масштаба.
Конечно, в реальной жизни нельзя бесконечно увеличивать масштаб, но фракталы по-прежнему являются важными математическими инструментами для понимания структур, которые самоподобны на разных масштабах, таких как реки и береговые линии.
Живые организмы демонстрируют очевидные фрактальные свойства в некоторых своих структурах, таких как ветви деревьев, бронхи и кровеносные системы. В таких структурах фрактальная геометрия на определённом диапазоне масштабов предлагает элегантное решение биофизической задачи. Например, кровь, несущая кислород и питательные вещества, должна достигать каждой клетки в организме, что означает, что оксигенированная кровь должна перекачиваться через разветвлённую сеть трубочек, которые, с точки зрения евклидовой геометрии, являются одномерными и должны многократно делиться, чтобы «заполнить» трёхмерное пространство. Таким образом, мы имеем фрактально разветвлённые (а затем сливающиеся) сети артерий, артериол, капилляров, венул и вен.
Изучив свойства дизайна таких фрактальных сетей и их последствия, теоретический физик Джеффри Уэст и его коллеги из Института Санта-Фе выяснили множество фундаментальных «законов масштабирования» в биологии.
Одним из самых известных результатов этой работы является объяснение того, почему каждое млекопитающее сердце будет биться около 1,5−2 миллиардов раз за свою жизнь, независимо от того, принадлежит ли оно этрусскому землеройке (весом в двадцатую часть унции и живущему около года) или синему киту (весом более трёхсот тысяч фунтов и живущему около века).
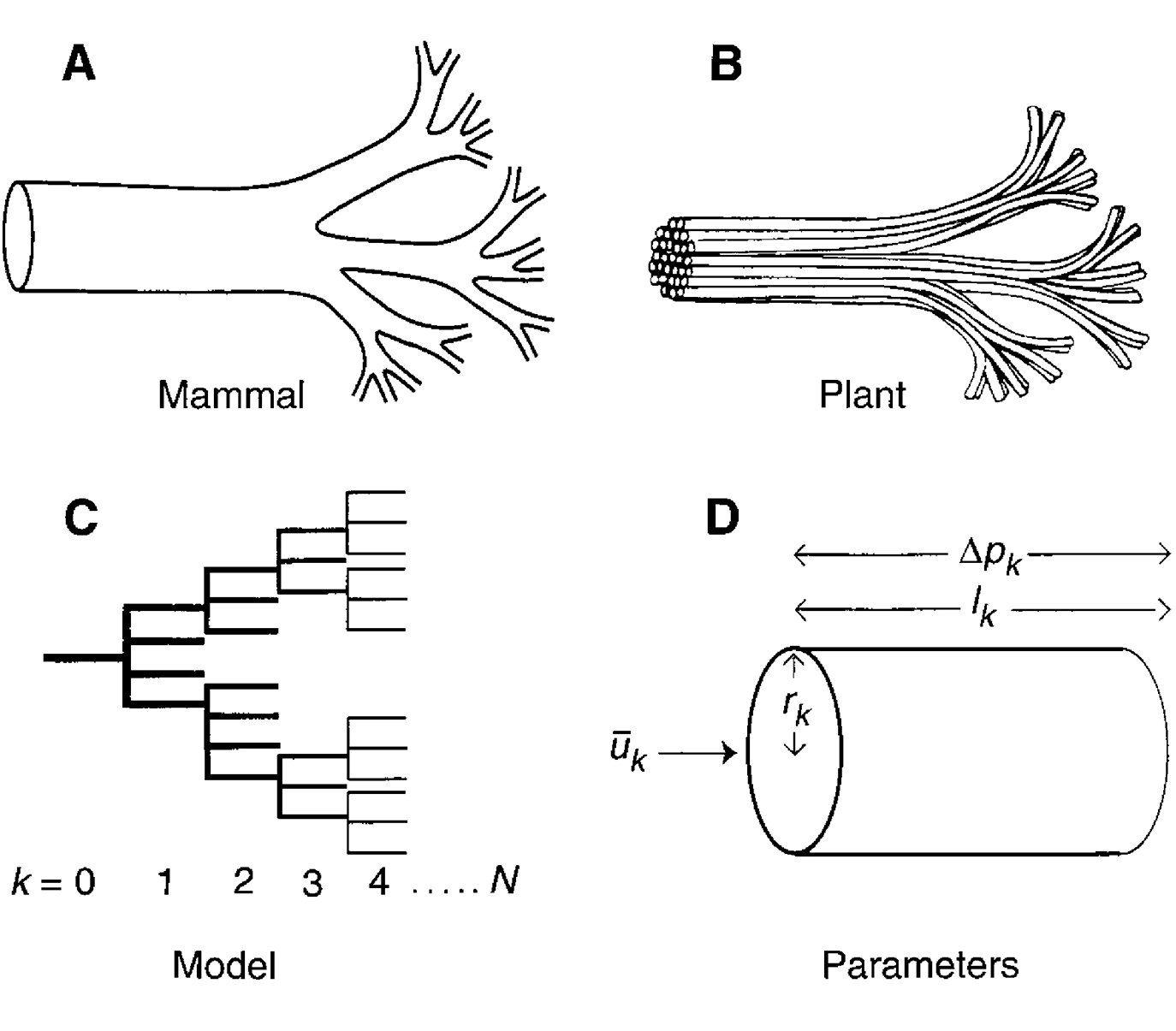
Фрактальное разветвление в кровеносных системах млекопитающих и растениях, по данным Уэста, Брауна и Энквиста, 1997 год.
Самоподобие в общем приводит к «законам степени» масштабирования, которые выглядят как прямые линии при построении на логарифмических осях.
Например, масса тела имеет законы степени в отношении метаболической активности, общей длины кровеносной системы и продолжительности жизни.
Сочетание различных степенных зависимостей для расчета количества сердечных сокращений за жизнь приводит к тому, что все масштабные переменные прекрасно взаимно уничтожаются, в результате чего мы получаем знаменитую константу в 1,5–2 миллиарда ударов. Хотя эта теория не является точной (например, люди живут несколько дольше, чем предсказывает модель), она удивительно хорошо объясняет широкий спектр явлений с минимальными затратами.
Так является ли жизнь в целом своего рода фракталом? Нет. В описании фракталов часто вспоминается рифма британского математика Огастуса де Моргана: У больших блох есть маленькие блохи на спинах, чтобы кусать их, А у маленьких блох есть ещё меньшие блохи, и так до бесконечности. Стихотворение забавное именно потому, что абсурдно: в отличие от идеализированного фрактального побережья, живые системы, как правило, глубоко различны на разных масштабах. Хотя существуют редкие примеры, о которых говорит де Морган — такие как гиперпаразитические осы, крошечные паразиты, откладывающие яйца внутри больших ос — в общем случае мы не увеличиваем масштаб, чтобы обнаружить меньшую блоху, не говоря уже о бесконечности.
На самом деле даже побережья лучше всего характеризовать в реальной жизни как «мультифракталы»: системы с повторяющейся структурой на каждом масштабе, но с изменчивостью между масштабами. В результате их степенные законы также могут варьироваться. Например, увеличение масштаба вдвое на одном уровне увеличения, где рельеф особенно выражен, может привести к тому, что видимая длина увеличится в 2 1.5 ≈2.83 раза, в то время как увеличение масштаба вдвое на уровне, где побережье выглядит более гладким, может привести к увеличению видимой длины всего на 2 1.16 ≈2.24 раза. Каждая неровность имеет неровности на себе, но неровности на одном масштабе отличаются по форме от неровностей на другом.
Аналогично, хотя геномы всегда реплицируются и состоят из других геномов, которые также реплицируются, каждое целое отличается от своих частей, так же как интегральная схема отличается от транзистора.
Как же можно количественно оценить это многофрактальное свойство в геноме? Ответ тесно связан с сжатием данных. Рассмотрим bff. В своем начальном состоянии ленты состоят из случайных байтов; здесь нет повторяющейся структуры. Если бы мы читали ленты по одному байту за раз, следующий байт имел бы равную вероятность принять любое значение от 0 до 255, независимо от предыдущих байтов. Таким образом, последовательность полностью непредсказуема, и если бы вы попытались сжать этот «суп» с помощью программного обеспечения для сжатия файлов, такого как ZIP, компрессор не смог бы уменьшить размер файла. (Это, возвращаясь к термодинамике, рабочее определение полностью неупорядоченного состояния.)
Данные являются избыточными— то есть, сжимаемыми— как раз в той степени, в какой они предсказуемы или, эквивалентно, демонстрируют порядок. Алгоритмы сжатия, такие как те, что используются в ZIP, работают, ища уже встреченные в потоке данных шаблоны и используя эти шаблоны для кодирования последующих данных с минимальным количеством битов. Обычный прием заключается в том, чтобы постепенно создавать словарь или «фразеологический словарь» из исходного материала и заменять части текста указателями на этот словарь, когда указатель можно закодировать меньшим количеством бит, чем оригинальный текст. Словарь, по сути, является предсказательной моделью, и поскольку компрессор строит эту модель постепенно в процессе сжатия, декомпрессор может поэтапно восстанавливать идентичную копию модели в процессе декомпрессии.
При отсутствии каких-либо предварительных знаний о том, как моделировать длинную последовательность, описывать ее с точки зрения ранее встречавшихся частей — хорошая стратегия. Полный текст этой книги сжимается до примерно тридцати пяти процентов от своего оригинального размера в виде сырой текстовой строки — поскольку некоторые буквы встречаются чаще других, многие слова повторяются, а эти слова часто объединяются в устойчивые фразы.
Когда bff резко переходит к полному репликационному процессу, ленты неожиданно становятся крайне сжимаемыми; они уменьшаются до всего лишь пяти процентов от своего первоначального размера. Интересно, что многие участки человеческого генома тоже обладают высокой сжимаемостью. Эти потоки данных так хорошо сжимаются, потому что в любой момент последовательности можно довольно точно угадать, какая буква будет следующей, просто найдя самую длинную предыдущую последовательность, которая соответствует текущему контексту.
Такие совпадения возникают из-за репликации. В человеческом геноме мы говорим не о репликации на уровне индивида, а на уровне транспозируемых элементов (или их ископаемых остатков) внутри нашего генома. Аналогично, даже одна лента bff обладает высокой сжимаемостью, потому что она образовалась в результате симбиотического слияния более мелких репликаторов… и эти репликаторы, скорее всего, являются либо копиями друг друга, либо родственными, образовавшись из еще более мелких несовершенных репликаторов… которые также могут быть копиями или родственными.
Как фракталы, последовательности символов, созданные путем многократного копирования подпоследовательностей всех длин, демонстрируют масштабирование по степенному закону — в данном случае, между длинами повторяющихся последовательностей и их частотой. Например, реплицированная последовательность в два раза длиннее другой может встречаться в два раза реже — что довольно часто! (Эта статистика была бы совершенно иной для случайных последовательностей, где увеличение длины последовательности привело бы к гораздо более быстрому экспоненциальному снижению ожидаемой частоты.) Последовательности, построенные согласно фиксированному правилу самокопирования, в определенном смысле бесконечно сжимаемы, поскольку они содержат лишь столько информации, сколько потребуется для записи рецепта, независимо от длины последовательности.
В отличие от этого, в сложной реплицирующей системе, такой как bff — не говоря уже о геноме — способ, которым любая конкретная группа реплицирующих подпоследовательностей объединяется для формирования более крупной реплицирующей последовательности, всегда будет специфичным.
Это будет зависеть от функциональных особенностей того, как эти части соединяются, чтобы создать работоспособное целое. Поэтому каждая такая комбинация добавляет новизну или информацию. Однако эта информация будет значительно меньше, чем необходимо для задания произвольной последовательности символов той же длины.
Когда мы ищем это свойство в bff и данных человеческого генома, мы именно это и находим! Оба они не только хорошо сжимаются, но и сжимаются всё лучше по мере увеличения объема данных. Например, одна лента bff сжимается в определенной степени, но вся «суп» сжимается лучше, и по мере роста объема «супа» коэффициент сжатия продолжает улучшаться — хотя, независимо от того, сколько уже было увидено, оставшаяся часть никогда не будет полностью предсказуема.
Аналогично, хотя отдельный участок человеческого генома уже хорошо сжимается, весь геном сжимается ещё лучше. В статистическом контексте множества других человеческих геномов так много информации в вашем геноме избыточна, что её можно сжать до размера файла, достаточно маленького, чтобы отправить его в виде вложения к электронному письму.
Эта закономерность продолжается, если мы посмотрим ещё шире и учтем геномы различных видов. Поэтому существует знаменитая статистика, что человеческий геном на девяносто восемь процентов идентичен геному шимпанзе… и на шестьдесят процентов похож на геном плодовой мухи!
Хотя общее предковство может объяснить многие сходства между видами (хотя не все, как иллюстрирует BovB), такие классические дарвиновские аргументы не могут объяснить, почему геном одного индивидуума также так повторяется. Это становится понятным только тогда, когда мы начинаем рассматривать геномы как состоящие из меньших геномов.
Другими словами, ДНК и ленты bff — это обе системы, которые эволюционируют через то, что мы могли бы назвать «мультифрактальной симбиозой». Наши геномы — это агрегаты сотрудничающих репликаторов, начиная с самого низа. Вот почему они так хорошо сжимаются.
Но именно поэтому на каждом уровне возникают удивительные новые виды сложности — и именно поэтому в биологии мы всегда можем узнать что-то новое, увеличивая или уменьшая масштаб.
Воплощение
Существует глубокая, но тонкая связь между мультифрактальными свойствами наших тел и мультифрактальными свойствами генома.
Скелет змеи из книги Лидеккера,
«Королевская естественная история», 1893
Эта связь тонка, потому что генетический код вовсе не является однозначным чертежом, явно представляющим окончательную форму тела. Можно было бы представить, что, поскольку у змеи сотни ребер, в ее ДНК может быть участок, кодирующий пару позвонков и ребер, который мог бы воспроизвестись несколько сотен раз в геноме змеи. Могут ли змеи быть результатом активного ретротранспозона, кодирующего ребра, подобно BovB?
Не совсем. Так называемые «гены Hox», широко распространенные среди животных, контролируют общий план тела и «реализуют» свою программу, используя комбинацию регуляции генов и распределенных вычислений, о которых Тьюринг говорил в своей работе по морфогенезу. В процессе эмбрионального развития ребра начинают как самоорганизованная стоячая волна в концентрациях химических морфогенов — тот же процесс, который придает коровам пятна, а тиграм — полосы. Однако в отличие от паттернов окраски кожи, которые просто связаны с изменением пигментации, программа развития ребра включает множество последующих морфогенетических этапов.
Этот механизм значительно мощнее, универсальнее и эволюционно изменяемее, чем простое воспроизведение какого-то кода «построить ребро». Он повторно использует код, как и любой достойный программист: вызывая одну и ту же функцию «построить ребро» сто раз вместо того, чтобы копировать и вставлять ее сто раз. Таким образом, изменение гибкости или кривизны ребра требует внесения одного изменения, а не ста.
 плодовой мухи приводит к тому, что из головы вместо антенн вырастают дополнительные ноги, что иллюстрирует композицией в построении тела.
Вверху: дикий тип;
Внизу: мутант.
Эволюция может создавать настоящие программы, которые повторно используют код, потому что жизнь является вычислительной. Помните, что если вы являетесь репликатором, о котором говорил фон Нейман, вам необходимо содержать инструкции для самостроительства, включая машину B для копирования этих инструкций и машину A, «универсального конструктора», для их выполнения. Универсальный конструктор — это движущая сила не только размножения, но и роста, развития, исцеления и поддержания. Он создал вашу сердечно-сосудистую систему и кожу в процессе эмбрионального развития; он также заживляет вашу кожу и восстанавливает разорванные капилляры, когда вы получаете порез. Пока вы живы, ваше тело будет непрерывно строить и перестраивать себя. Код никогда не останавливается.
И поскольку универсальный конструктор является своего рода универсальной машиной Тьюринга, его вычисления, или, что эквивалентно, математические функции, по своей природе «композиционны»: сложные функции (или программы) могут состоять из подфункций, которые, в свою очередь, могут состоять из под-подфункций… пока, в конечном итоге, всё не может быть выражено через элементарные инструкции машины Тьюринга.
Функции могут даже состоять из
себя
. В информатике это известно как «рекурсия». Последовательность Фибоначчи, например — 1, 1, 2, 3, 5, 8, 13, 21 и так далее, где каждое число является суммой двух предыдущих — проще всего выразить как рекурсивную функцию f(n) = f(n−1) + f(n−2) для чисел больше 1, с f(0) = f(1) = 1. Мы находим числа Фибоначчи в биологии постоянно, как, например, в спиральном узоре шишки сосны, головке подсолнечника и камерах наутилуса. Это указывает на то, что рекурсивные вычисления происходят в построении сосен, подсолнечников и наутилусов.

1896 год, skiagraph (рентгеновская фотография)
Nautilus pompilius
Однако настоящим доказательством является рост самоподобных структур в организме. Композициональность и рекурсия позволяют генетическому коду строить многоуровневый фрактал, подобный кровеносной системе. Код для построения рёбер или сегментов наутилуса, теоретически, можно было бы просто скопировать несколько сотен раз, но это невозможно для артерий, вен или капилляров, где количество сегментов и ветвей становится астрономически большим. Код ветвления кровеносных сосудов
должен
переиспользоваться. И с относительно незначительными изменениями, должно быть возможно настроить параметры этого кода так, чтобы ветвление прекращалось на соответствующем размере для землеройки или продолжалось, создавая что-то размером с синего кита, поскольку землеройки и киты, в большом масштабе, являются близкими родственниками.
С эволюционной точки зрения композициональность подразумевает иерархическую сеть симбиотических отношений. Бактерии, ставшие митохондриями, и археи, которые их поглотили, изначально обладали кодом, необходимым для самовоспроизводства. Когда они объединились в эукариот, им всё ещё нужно было сохранить инструкции для собственного размножения, но также необходимо было развить дополнительные функции для взаимной регуляции. Когда эукариоты стали многоклеточными, тот же процесс должен был повториться; наши клетки всё ещё знают, как воспроизводиться индивидуально, и, действительно, клеточное размножение является фундаментальной операцией, вовлечённой в более крупномасштабный рост и размножение всего организма.
То, что верно для эволюции, также верно и для
развития отдельного организма.
Кровеносные сосуды, например, не просто евклидовы отрезки, а трубки, состоящие из слоёв гладкомышечных клеток. Каждая из этих клеток содержит набор органелл, а каждая митохондрия в каждой из этих клеток содержит свои собственные мембраны и кольцо бактериальной ДНК.
Живой организм — это композиционная структура в высшей степени . Он мог быть создан только вычислительно, через сочетание множества функций. И жизнь могла развиваться лишь как иерархическая композиция этих функций — то есть, через симбиогенез.
Дейзиворлд
Все определения жизни вызывают споры, большинство из них требует оговорок или исключений, и ни одно не продвинуло наше понимание биологии. Разочарованный таким положением дел, биохимик Эндрю Эллингтон даже утверждает, что «жизни не существует. Жизнь — это незначительный научный термин. Это термин для поэтов, а не ученых».
Позиция Эллингтона может показаться радикальной, но она более обоснованна, чем догматичное определение во многих учебниках биологии: живые организмы обладают «способностью есть, метаболизировать, выделять, дышать, двигаться, расти, размножаться и реагировать на внешние стимулы».
По этому определению вирусы не считаются живыми, что может показаться разумным. Но и муулы, и рабочие пчелы тоже не живы, поскольку они стерильны!
Или рассмотрим более лаконичное определение, популярное среди астробиологов: «жизнь — это самообеспечивающаяся химическая система, способная к дарвиновской эволюции».
Лучше, но по этому определению паразиты не считаются живыми, поскольку они не являются самообеспечивающимися. С другой стороны, есть ли хоть одна жизненная форма, которая действительно самообеспечивающаяся? (Не если она питается.) Как можно осмысленно определить «я»? И хотя жизнь на Земле основана на химии, стоит ли нам настаивать на этом как на требовании? Функционализм утверждал бы иное.
Итак, вот моя собственная попытка: Жизнь — это самоизменяющийся вычислительный материал, возникающий в результате отбора на динамическую стабильность; она эволюционирует через симбиотическую композицию более простых динамически стабильных сущностей.
Обратите внимание, что в моем определении не указана никакая конкретная функция. Тем не менее, определяя жизнь вычислительно, я подчеркиваю ее функциональность.
И в мире, где действует Второй закон, динамическая стабильность подразумевает, что эти функции должны включать в себя какую-то комбинацию самостроительства, самовосстановления или воспроизводства. Все это — вычисления, требующие наличия кода, способного читать и записывать код, отсюда и термин «самомодифицирующийся».

М. К. Эшера,
«Рептилии»,
март 1943
«Компьютрониум» обычно понимается как фаза материи, но под «материей» здесь может подразумеваться все, что может поддерживать вычисления, включая байты в bff или пиксели в клеточном автомате. Более того, в отличие от газов, жидкостей или твердых тел, которые в основном однородны на макроскопическом уровне, фаза материи, связанная с жизнью, неизбежно имеет неоднородную структуру. Одна из причин заключается в том, что однородный материал не может выполнять общие вычисления; компьютрониум должен, на своем самом мелком уровне, содержать различные взаимодействующие компоненты.
Существует и более глубокая причина. Динамическая стабильность подразумевает существование дискретных единиц — «я» или популяции «я». Без какой-либо модели «я» или «я» невозможно было бы сказать, что является стабильным. А поскольку жизнь формируется симбиотически, существуют «что-то» на множестве уровней — в нашем случае от молекул до митохондрий, клеток, целых организмов и далее. Изучение каждого «что-то» требует изменения масштаба, чтобы сосредоточиться на динамической стабильности сущности другого типа.
На самом широком масштабе жизнь представляет собой единую «кучу»; для нас это была бы Земля. Является ли она тоже живой? Физик и астробиолог Сара Уокер утверждает, что нет, именно потому, что она является единственным экземпляром. Уокер считает, что Земля станет живой только тогда, когда мы колонизируем другую планету, тем самым «воспроизводя» себя.
Мое определение жизни не требует воспроизводства, но требует динамической стабильности; можно ли это вообще определить для единственного экземпляра? В определенном смысле это возможно, и впервые это было формализовано в 1970-х и 1980-х годах независимым ученым Джеймсом Лавлоком вместе с Линн Маргулис и другими.
Их «гипотеза Гайи», названная в честь греческой богини Земли, основана на наблюдении, что наша планета в целом обладает гомеостатическими свойствами. Отрицательные обратные связи активно поддерживают различные параметры биосферы в пределах, благоприятных для жизни (хотя многие механизмы, такие как тот, что регулирует соленость морей, все еще плохо изучены). Например, температура Земли колебалась в относительно узком диапазоне на протяжении сотен миллионов лет, несмотря на то, что Солнце со временем становилось горячее. Температура Земли остается достаточно стабильной для жизни, потому что она активно согревает и охлаждает себя, и делает это потому, что она живая.
Механизм, благодаря которому возникает такая саморегуляция, не является загадкой и, по сути, сводится к дарвиновскому отбору для динамической стабильности. Модель сильно упрощенной планеты, которую Лавлок назвал «Дейзиуорлд», иллюстрирует, как это работает.
В Дейзиуорлде всего два вида: черные и белые ромашки, репродуктивные способности которых зависят от температуры — оптимальная температура составляет 22,5°C, постепенно снижаясь до нуля, если тепло ниже 5°C или выше 40°C. Черные ромашки согревают поверхность планеты, поглощая солнечный свет, в то время как белые ромашки охлаждают ее, отражая солнечные лучи. Тщательный математический анализ показывает, что популяции черных и белых ромашек будут динамически регулировать среднюю температуру поверхности, поддерживая ее близкой к оптимальной в диапазоне солнечной светимости — замечательный диапазон, который на безжизненной планете привел бы к температурам от 0°C до 60°C.
Популяции черных и белых ромашек работают в тандеме, чтобы стабилизировать температуру в Дейзиуорлде в диапазоне солнечной светимости; A. J. Watson и Лавлок, 1983.
Хотя детальный анализ сложен, даже для простой экосистемы из двух видов, механизм, стоящий за этой гомеостатической тенденцией, является универсальным.
Грубо говоря, это утверждение верно, когда существуют виды, популяции которых зависят от окружающей среды и одновременно влияют на неё. Некоторая гомеостазис наблюдается даже при наличии всего одного вида (в мире ромашек это могут быть либо черные, либо белые ромашки), но гомеостазис становится более устойчивым с несколькими видами, которые, подобно противоположным мышцам, могут изменять окружающую среду в ту или иную сторону.
Таким образом, на мой взгляд, планета, такая как мир ромашек или Земля, действительно жива, поскольку она не только содержит вычислительные формы жизни (и, следовательно, в совокупности выполняет множество вычислений), но и саморегулируется для достижения динамической стабильности. Другими словами, мир ромашек демонстрирует, как эволюция может продолжать действовать даже при размере популяции в один экземпляр. Ключевым моментом является модульность: её части также живы и могут подвергаться эволюционному отбору.
Леса осин предлагают полезную параллель. Осиновые деревья пускают горизонтальные корни, которые могут прорастать в ствол и превращаться в другое дерево. Или это другой ствол того же дерева? «Пандо», расположенный в округе Севьер, штат Юта, представляет собой трясущуюся осину, занимающую более ста акров и весом, по оценкам, более тринадцати миллионов фунтов. Хотя каждое из его сорока семи тысяч деревьев (стволов? стеблей?) имеет конечный срок жизни, весь лес можно рассматривать как одно бессмертное существо, которое, если бы оно находилось в плодородной долине, окруженной непроходимыми горами, возможно, никогда бы не размножалось.
▶ Трясущаяся осина «Пандо», штат Юта
Хотя генетика осинового леса заперта в рамках — поскольку каждое видимое дерево является клоном — дарвиновский отбор все же происходит: стволы в благоприятных местах будут процветать и размножаться, в то время как те, что находятся в неблагоприятных условиях, не смогут. Индивидуальные деревья не могут двигаться, но лес может — это пример «возникающей способности». По мере изменения климата в долине весь лес начинает двигаться, поднимаясь и опускаясь по склонам гор для поддержания гомеостаза.
Таким образом, вполне разумно называть как деревья, так и лес живыми, так же как разумно считать живыми и вас, и ваши отдельные клетки. То же самое можно сказать и о Земле в целом. Однако это не означает, что Земля «умнее» суммы своих частей или что она обладает сознанием только потому, что мы, будучи её частью, обладаем им. На самом деле гипотеза социальной интеллигентности, которую мы рассмотрим в главе 5, предполагает, что наше сознание возникло в результате социальных взаимодействий. У нашей планеты нет других живых планет, с которыми можно было бы общаться.
Земля обладает интеллектом лишь в той мере, в какой модели, заложенные в её обратных связях, позволили ей выжить в условиях постепенных изменений на протяжении миллиардов лет — это медленный, вегетативный вид интеллекта с ограниченной предсказательной силой. Хотя сейчас техногуманность начинает моделировать как Землю, так и небеса, держа телескопические «глаза» открытыми для блуждающих астероидов и радио «уши» настороженными для внеземных сигналов. Пока безрезультатно… Но можно утверждать, что наша планета стремительно становится более интеллектуальной и «животной», способной чувствовать удалённо и делать долгосрочные прогнозы, готовой проактивно реагировать на угрозы и возможности, если они возникнут — при условии, что мы сможем организоваться и восстановить планетарную гомеостазу, включающую человечество и технологии.
Élan Vital
Если жизнь является вычислительным процессом, то являются ли компьютеры, созданные человеком (то есть «искусственные»), живыми? Они, безусловно, имеют цель, иначе мы не могли бы говорить о них как о сломанных или с ошибками. Но аппаратное и программное обеспечение, как правило, не способны воспроизводиться, расти, исцеляться или эволюционировать самостоятельно, поскольку инженеры давно поняли, что саморефлексирующий код (как bff или ДНК) трудно понять и отладить.
Таким образом, телефоны не производят «детей», и, увы, не исцеляются от своих частых травм. Приложения не пишут новые версии самих себя.
И всё же: в этом году в мире больше телефонов, чем в прошлом, и их треснувшие экраны действительно часто ремонтируют; приложения получают новые функции, устаревают и в конечном итоге выходят из употребления, уступая место новым; модели ИИ улучшаются с каждым месяцем. Электронные компоненты и компьютерный код также демонстрируют ту же композиционность, которую мы наблюдали в bff и ДНК. Безусловно, кажется, что технологии воспроизводятся и эволюционируют! Дебаты о том, живы ли они, напоминают споры о том, живы ли вирусы или паразиты.
▶ Отрывок из Королевства фей Жоржа Мелиеса, 1903
Если мы отдалимся и поместим технологии и людей в один кадр, то увидим, что это более широкое симбиотическое «мы» действительно воспроизводится, растёт и эволюционирует. Появление технологий и взаимовыгодные — хотя иногда и напряжённые — отношения между людьми и технологиями не что иное, как наш последний крупный эволюционный переход. Таким образом, технологии не отделены от природы или биологии, а являются лишь их последним эволюционным развитием.
А что насчёт тех старинных неживых вещей — камней и рек, гор и пляжей, облаков и бурь? Молекулы воды сами по себе явно не способны к общей обработке информации. Тем не менее, в контексте гидрологического цикла облака, дожди и реки, безусловно, выполняют критически важные экологические функции и глубоко формируются жизнью. Точно так же металл и песок нашей планеты превращаются в паровые машины и компьютерные чипы, как кальций, который мы усваиваем, становится костями и ионными токами. Все эти материалы и структуры составляют части великой сети Гаевой взаимозависимости.
Почему мы проводим границы вокруг определенных сетей функций и настаиваем на том, что они «живы», утверждая при этом, что окружающие функции не являются таковыми?
Такой подход ведет к витализм, взгляду, который в различных формах поддерживался долгой чередой философов — от Посидония Апамейского (около 135–51 гг. до н. э., и, безусловно, отражающего гораздо более древнюю традицию) до Анри Бергсона (1859–1941). Некоторые современные мыслители также защищают виталистскую позицию, например, Джейн Беннет:
Карантины материи и жизни побуждают нас игнорировать жизненную силу материи и живые способности материальных форм […]. Под жизненной силой я подразумеваю способность вещей — съедобных, товаров, бурь, металлов — не только препятствовать или блокировать волю и замыслы людей, но и действовать как квази-агенты или силы с собственными траекториями […] или тенденциями. [… Наши] анализы политических событий могут измениться, если мы уделим больше внимания силе вещей.
Мы сопротивляемся таким идеям, потому что склонны резервировать понятие агентности только для себя. Идея агентности в молекуле или буре, не говоря уже о такой абстракции, как деньги, кажется особенно надуманной. Мы также склонны мыслить в терминах иерархии, в которой «мы» (при любом значении «мы») находимся на вершине, и агентность, безусловно, должна уменьшаться для всего «низшего» — взгляд, напоминающий средневековую Великую Цепь Существования.
Когда мы (с колебаниями) расширяем понятие «агентности» на нечеловеческое, мы, как правило, делаем это только для вещей, которые действуют явно, индивидуально и в быстрых временных масштабах, а не в совокупности и на более медленных, эволюционных. Возможно, пришло время пересмотреть эти идеи более целостно.
Моя цель здесь не следовать по стопам Беннет — хотя я считаю ее проект достойным серьезного внимания. Язык, по своей сути, неточен, независимо от того, какие определения мы выбираем. Это не означает, что слова бесполезны.
Когда наш язык становится более строгим и научно обоснованным, и когда мы используем его для описания последовательных паттернов, объединяющих широкий спектр наблюдений, мы начинаем видеть сквозь идеологические заросли.
Я надеюсь, что смог ясно и строго объяснить, как явления, порождающие сложные динамики жизни, применимы гораздо шире, чем к тем сущностям, которые мы обычно считаем «живыми» или «агентными». Соответственно, мы могли бы расширить наши определения этих существующих слов, или принять новые, или сделать немного того и другого. Лично я считаю, что расширение старых повседневных слов было бы полезным.
Это вряд ли откроет новые горизонты. Многие традиционные, номинально «преднаучные» мировоззрения охватывают понятия живости, агентности и даже личностности, которые гораздо шире современных западных. Это, похоже, яркий пример сходящейся эволюции языков и идей, вызванный общей необходимостью традиционных обществ серьезно воспринимать симбиоз для обеспечения собственного выживания и процветания. Более широкий взгляд практичен так же, как и духовен: поощрение более богатого моделирования агентности в «природе» усиливает динамическую стабильность общества, поскольку все в «природе», включая нас самих, взаимозависимы.

Рисунки аборигенов Аṉaŋu, Национальный парк Улуру-Ката Тьюта, Австралия
Таким образом, иногда полезно взглянуть на мир глазами животного, растения или реки, даже если они не могут взглянуть на мир нашими глазами, чтобы лучше заботиться о них — и о себе. То же самое касается Земли в целом. В конечном итоге, это лучшая причина рассмотреть возможность принятия, или хотя бы адаптации, более инклюзивного взгляда на живое. Писательница и биолог Потаватоми Робин Уолл Киммерер убедительно излагает эту мысль в своей книге «Заплетая сладкую траву».
Согласившись с Киммерер, я не призываю к суеверию.
Когда ученые осуждают анимистические верования как суеверные, они обычно ссылаются на материалистические открытия Просвещения, которые показывают, что атомы, из которых состоят наши тела, не отличаются от атомов, из которых состоят камни или воздух. Это правда. Атомы — это атомы; они все подчиняются одним и тем же законам. Следовательно, модель вселенной как часового механизма, управляемого динамическими законами, характерна для эпохи Просвещения.
Тем не менее, как указал Шрёдингер в 1944 году, наше понимание этих законов — в разработке которых он сыграл центральную роль — остается неполным. Законы, как они есть, не учитывают вычислительно сложные, динамически стабильные, симбиотические явления, которые составляют большую часть нашего опыта на Земле — без которых, по сути, не было бы никакого опыта вообще. Не было бы жизни, цели, разума или воли.
Когда мы как следует принимаем научную строгость и начинаем разбираться в тех «‘других законах физики’, которые до сих пор неизвестны», возможно, нам стоит меньше удивляться, обнаруживая, что ситуация обернулась иначе. То, что поэт Дилан Томас назвал «силой, что через зеленую нить ведет цветок», движет всеми атомами, а не только теми, которые мы предполагаем быть живыми.

Деталь картины Ялти Напангати, изображающая Маррапинти, Западная пустыня, Австралия
W. Thomson 1871
↩
.
Перетó, Бада и Ласкано 2009
↩
.
Робертсон и Джойс 2012
↩
.
М. Дж. Рассел и Мартин 2004; Шапиро 2006
↩
,
↩
.
Помните из урока химии, что поскольку водород (H) состоит из одного протона, связанного с одним электроном, положительно заряженный водородный ион, у которого электрон отнят (H+), — это просто протон. Когда концентрация протонов варьируется в пространстве, возникающий поток ионов генерирует электрический ток.
Кребс и Джонсон 1937
↩
. Кребс получил Нобелевскую премию по медицине за эту работу в 1953 году.
М. С. Эванс, Бьюкенен и Арнон 1966
↩
.
Бьюкенен и Арнон 1990
↩
.
Сегодня подавляющее большинство органического топлива, потребляемого дыханием у животных и грибов, происходит от растений, которые фиксируют углерод с помощью фотосинтеза, а не более древнего обратного цикла Кребса.
Szathmáry и Smith 1995
↩
. Более узкий термин «эволюционные переходы в индивидуальности» (ETI) также становится все более популярным; Michod 2000
↩
.
Sagan 1967
↩
. Woese 2002
↩
. В этой книге я буду использовать термины
симбиоз
и
симбиогенез
в более широком смысле, чем это делают большинство биологов, чтобы включить в них сотрудничество и последующую взаимозависимость между сущностями всех видов — в духе молекулярных биологов, которые рассматривают происхождение жизни как симбиоз между «молекулами в взаимовыгодных отношениях», согласно Lanier, Petrov и Williams 2017
↩
. См. также Mizuuchi, Furubayashi и Ichihashi 2022
↩
. Konstantinidis, Ramette и Tiedje 2006; Murray, Gao и Wu 2021
↩
. Raup и Gould 1974
↩
. В некоторых случаях, как, например, автомобиль против лошади с повозкой или фотосинтез против обратного цикла Кребса, новая технология вытесняет старую благодаря большей эффективности; однако даже здесь вытеснение часто не является полным. Все еще остаются несколько лошадей и повозок, а также некоторые организмы, использующие обратный цикл Кребса.
Church 1936; Turing 1936
↩
. Turing 1937
↩
. Технически машина Тьюринга, будь она универсальной или нет, всегда должна иметь возможность увеличивать размер своей ленты при необходимости, в то время как реальные компьютеры имеют фиксированные ограничения по объему памяти — но для большинства целей этого достаточно.
von Neumann 1945
↩
. Оба они умерли молодыми: Тьюринг в 1954 году в возрасте сорока одного года, а фон Нейман в 1957 году в возрасте пятидесяти трех лет.
Turing 1952
↩
. Много лет спустя немецкий молекулярный биолог Кристиане Нюссlein-Вольхард идентифицировала первый реальный морфоген — белок «Бикоид», который формирует градиент вдоль эмбриона плодовой мушки, чтобы установить его сегментированный план тела. За это открытие она получила Нобелевскую премию вместе с Эриком Висхаусом и Эдвардом Льюисом в 1995 году.
von Neumann и Burks 1966
↩
.
В более абстрактной форме математик Стивен Клейн доказал этот результат много лет назад, который сегодня известен как Вторая теорема о рекурсии Клейна; Клейн 1938.
Я использую термин «сложный» здесь в разговорном смысле; согласно как минимум одной технической дефиниции (комплексность Колмогорова), если инструкции внутри «простого» объекта полностью описывают, как создать «более сложный», они на самом деле имеют одинаковую сложность. Д. Д. Уотсон и Фрэнсис Крик 1953.
Хотя обратные вычисления теоретически возможны и в последние годы привлекли renewed интерес из-за своей связи с квантовыми вычислениями, они остаются в значительной степени неизученными. Тьюринг 2000 [1951].
Тьюринг 1948. Мозги младенцев, конечно, не случайны. Тьюринг был прав, полагая, что случайность играет важную роль в их первоначальной «схемотехнике»; последующее обучение и развитие укрепляют некоторые связи, в то время как многие другие отсекаются. Песавенто 1995.
Мордвинцев и др. 2020. Агера и Аркас и др. 2024. Эта работа черпает вдохновение из классических исследований в области ALife, особенно из работ Баррикелли 1957; Фонтана 1990; Рея 1991; Адами и Брауна 1994.
Уменьшение 0 оборачивается к 255, максимальному возможному значению для байта, а увеличение 255 оборачивается к 0. Вы, возможно, заметили, что оригинальный Brainfuck имел восемь инструкций, включая « . » для «печати» и « , » для «ввода». Bff использует только « , », потому что вместо чтения и записи на внешний терминал или консоль лента читает и записывает на саму себя . Таким образом, копирование байта из одного места в другое требует всего лишь одной инструкции. В нашей первой статье, описывающей bff, мы включили инструкции « { » и « } » для перемещения указателя консоли, но упрощённая альтернатива, используемая здесь, просто инициализирует указатели данных и консоли первыми двумя байтами на ленте. Фонтана 1990.
Амато 1991. Шрёдингер 1944. Х. Рейхенбах 1956; Ровелли 2018.
Квалификатор «почти» возникает из глубокой математической структуры релятивистской квантовой механики. Ряд близких симметрий в физике, таких как положительный или отрицательный заряд, левосторонность или правосторонность, а также обращение времени, не являются абсолютно точными в квантовой полевой теории, хотя, согласно теореме CPT — буквы обозначают заряд, паритет и время — одновременное обращение всех трех величин действительно приводит к точной симметрии.
На данный момент я уклоняюсь от прямого определения термина «беспорядок» и его очевидной субъективности (является ли мой стол беспорядочным или он отражает порядок, который вижу только я?), но вскоре мы вернемся к центральной роли модели в различении порядка и беспорядка; это связь, соединяющая термодинамику с теорией информации.
Развитие часов и других сложных машин в Средние века создало фон, согласно Пригожину и Стенгерс 1984: «Мир часов — это метафора, намекающая на Бога-Часовщика, рационального мастера роботоподобной природы». Ньютоновская динамика усилила представление о том, что динамика вселенной детерминирована и познаваема, следовательно, «роботоподобна».
На данный момент я буду использовать термин «Индустриальная революция» для обозначения первой Индустриальной революции, начавшейся с использования ископаемого топлива примерно с 1800 года. Последующие революции, связанные с электрификацией (около 1900 года), компьютеризацией (около 1980 года) и ИИ (около 2020 года), будут рассмотрены в главе «Уязвимость», глава 10.
PV = nRT, где P — давление, V — объем, T — температура, n — количество молекул газа или шаров для бильярда, а R — «константа идеального газа». Кинетическая энергия пропорциональна массе и квадрату скорости молекул. Более точно, это отталкивающие взаимодействия между электронными оболочками молекул.
Это прекрасно иллюстрируется экспериментом 2024 года, связанным с «миром РНК», в котором в лабораторных условиях мутирующая, несовершенно реплицирующаяся молекула РНК эволюционирует в несколько форм, способных существовать бесконечно; Папаставру, Хорнинг и Джойс 2024.
В языке теории динамических систем термодинамическая стабильность является фиксированной точкой, в то время как DKS представляет собой предельный цикл. Доукинс 1986.
Кауфман 1971.
В классической мифологии химеры — это «неестественные» комбинации животных — например, слияние льва, козы и змеи. Как мы увидим, на самом деле они вовсе не так уж неестественны. Йинуса и Неханив 2011.
Это не абсолютное правило, поскольку новые части также могут эволюционировать внутри существующего целого. Ландер и др. 2001.
Балтимор 1970.
МакКлинток 1950.
Тарлингтон, Мерс и Янг 2006.
Ши и др. 2022.
Хорита и Фукумото 2023.
Чуонг 2018; Бакулис и др. 2022; Русс и Иорданский 2023.
Выключение ретротранспозона «Arc», вероятно, вирусного происхождения, делает лабораторных мышей неспособными формировать новые долгосрочные воспоминания, согласно Пастузын и др. 2018.
Райан 2009.
Шарп и Хан 2011.
Кросби 2003; Харпер и др. 2008.
Фешотт и Гилберт 2012; Навиль и др. 2016.
Насир и Каетано-Анолес 2015.
Ёнг 2013.
В отличие от эукариотов, прокариоты (бактерии и археи) имеют очень низкие пропорции «мусорной ДНК». Это связано с тем, что копирование и экспрессия ДНК требуют значительных энергетических затрат, а прокариоты ограничены в энергии, что ограничивает максимальную длину их ДНК и накладывает сильное эволюционное давление на избавление от кода, который не может выполнять свою функцию. Однако «основной геном» прокариота обычно составляет лишь 60–70 процентов от общего объема, в то время как оставшаяся часть сильно варьируется среди штаммов. (Таким образом, два E.
коли
бактерии могут отличаться друг от друга так же, как человек от плодовой мухи!) Горизонтальный перенос генов через транспозоны, вирусы или «плазмиды» (обменные петли ДНК) позволяет широко копировать фрагменты кода между бактериями, что приводит к сложному переплетению воспроизводящихся кусочков кода, концептуально схожему с тем, что описано здесь для эукариотов.
Хотя ранее мы также рассматривали отдельные инструкции bff как минимальные или «атомные» воспроизводящие единицы.
Мандельброт заново открыл и обобщил эти выводы в 1960-х; см. Мандельброт 1967
↩
.
Уэст 2017
↩
.
Для математически настроенных: степенные законы, связывающие две переменные
x
и
y
, принимают общий вид
y
=
ax
b
.
Уэст, Браун и Энквист 1997
↩
.
Де Морган и Де Морган 1872
↩
.
Уолперт и Макариди 2007; Багров и др. 2020
↩
,
↩
.
Это имеет смысл, поскольку процессы, формирующие береговые линии, различаются в зависимости от масштаба, скажем, метра и километра; см. Мандельброт 1989
↩
.
Эти конкретные числа соответствуют самым гладким,
N
=5, и самым грубым,
N
=8, диаграммам береговой линии Мандельброта из его классической статьи 1967 года «Какова длина побережья Британии?»
Если бы я был более изысканным писателем и использовал меньше стандартных фраз, книга не была бы такой эффективной. Используя тот же алгоритм сжатия, «Улисс» Джеймса Джойса можно сжать только до сорока процентов его исходного размера!
В математике: для случайной строки символов, которая может принимать
K
значений, строка длины
L
будет встречаться с частотой
f=K
–L
, в то время как при репликации на всех масштабах частота строки масштабируется как
f~L
–b
.
Кристли и др. 2009
↩
.
Рекурсия не может быть реализована рибосомой самостоятельно; я немного пофантазировал, идентифицируя рибосому с машиной A. На самом деле, как иллюстрирует морфогенез, биологические вычисления более целостны и распределены. Как «пишущая рука» репликатора фон Неймана, рибосома является лишь одной функциональной частью более крупной, универсальной биологической машины A.
Кэрролл, Гренье и Уэзерби 2013
↩
.
Эллингтон 2010
↩
Уокер 2024
↩
Уокер 2024
↩
Личное сообщение во время студии «Когнитивные инфраструктуры Антикиферы» в Центральном Сент-Мартинсе в Лондоне, июль 2024.
А. Дж. Уотсон и Лавлок 1983
↩
Как будет обсуждено в главе 5 «Экипаж из восьми», осьминог может служить примером более модульного пути к высокому интеллекту.
Компьютерные вирусы — это заметное исключение.
Аргумент о том, что технологии являются эволюционным продуктом иерархически составленных функций, тщательно обоснован с множеством примеров Артуром 2010
↩
Как сказал астроном Роберт Ястроб, «Мы — репродуктивные органы компьютера» (Ястроб 1981
↩
); то же самое справедливо и для технологий в целом. Самуэль Батлер изложил эту мысль в своем пророческом эссе 1863 года под названием «Дарвин среди машин», эта перспектива была переосмыслена и обновлена в конце двадцатого века историком науки Джорджем Дайсоном; Батлер 1863; Г. Дайсон 1998
↩
↩
Д. Беннет 2010
↩
Лавджой 1936
↩
Энфилд 2024
↩
Киммерер 2013
↩
Шрёдингер 1944
↩
Томас 1934
↩
Выживание